

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
There is growing evidence that as well as being the building blocks of proteins and polypeptides, some amino acids have roles in regulating key metabolic pathways that are necessary for maintenance, growth, reproduction, and immunity (1). They are called functional amino acids, which include arginine (Arg), cystein (Cys), glutamine (Gln), leucine (Leu), proline (Pro), and tryptophan (Try). Dietary supplementation with one or a mixture of these amino acids may be beneficial for ameliorating health problems at various stages of the life cycle, and for optimizing the efficiency of metabolic functions to enhance muscle growth, milk production, and athletic performance (1-5). Indeed, microarray analysis indicates that dietary supplementation with Gln or Arg increases the expression of anti-oxidative genes, and reduces the expression of pro-inflammatory genes in the small intestine and adipose tissues (6-8). In addition, some amino acids, such as Gln, Arg, and Leu are known to stimulate the phosphorylation of mTOR1 in a cell-specific manner, thereby regulating intracellular protein turnover (9-12).
Protein deficiency has long been known to impair immune function and increase the susceptibility of animals to disease. Some amino acids, including Gln, Arg, methionine (Met), have been recognized as playing a role in the enhancement of immune function (13-15). The underlying mechanism may involve mTOR activation, NO and glutathione synthesis, H2S signaling, and cellular redox state. In particular, catabolism of Try via indoleamine 2,3-dioxygenase (IDO) appears to be critical for functions of both macrophages and lymphocytes (16). Thus, anthranilic acid (a metabolite of tryptophan via the IDO pathway) inhibits production of proinflammatory cytokines and prevents autoimmune inflammation (17). Gln is a non-essential amino acid which is also considered as an essential nutrient during multiple injury, trauma, and tumor (18-22). Cell culture studies demonstrate that failure to supplement culture media with Gln impairs the ability of lymphocytes to respond to mitogenic stimulation (23). In terminally differentiated macrophages, Gln may be required for the synthesis of mRNA for producing secretory proteins in immune challenge during pinocytosis or phagocytosis (24). In addition, Gln has been shown to be beneficial in the prevention of infectious morbidity and mortality in seriously ill patients, due to its ability to maintain the integrity of the intestinal mucosal epithelium (25). It has also been suggested that Gln influences the growth of immune cells, T helper function and responsiveness, and synthesis of immunoglobulin A (IgA) (21,22,26). In support, inadequate concentrations of Gln have been shown to compromise lymphocyte proliferation (27), IL-2 and IFN-γ production (27,28), the expression of surface activation markers (CD25, CD45RO, and CD71) (28), and lymphokine-activated killer cell activity (29), and to decrease the proportion of CD4+ lymphocytes and the CD4+/CD8+ ratio (30). Leu, one of branched chain amino acids, which also include isoleucine and valine, is an essential amino acid, and its concentration increases in skeletal muscle but decreases in plasma. It was assumed that Leu is a possible regulator of protein turnover in muscle and the perfused liver. Also, Leu stimulates muscle protein synthesis under both in vitro and in vivo experimental conditions (31). The underlying mechanisms may involve activation of mTOR signaling to enhance translation initiation and to inhibit autophagy in liver and muscle (25). Interestingly, other branched amino acids, isoleucine and valine, have no effect on mTOR phosphorylation or muscle protein turnover (9,10), indicating a structural specificity of Leu in cell signaling and function.
Herpes simplex virus type 1 (HSV-1) is human pathogenic viruses, which can replicate in cells of the epithelial lineage, and cause mucocutaneous lesion, as manifested in orofacial and genital herpes (32,33). Importantly, HSV-1 is also neurotropic viruses, and establish latent infection in neurons, which can be reservoirs of virus during subsequent events of reactivation. The morbidity and socioeconomic burden associated with genital herpes as well as the alarming relationship between genital herpes and the increased risk of acquiring a HIV infection emphasize the need for development of an effective vaccine and/or therapeutics (32,33). Based on the accumulated results of biological functions of amino acid, we evaluated the antiviral effect of several amino acids against mucosal infection of HSV-1 in the present study. We found that Gln and Leu have specific roles in regulating protective immunity against mucosal infection of HSV-1.
MATERIALS AND METHODS
Mice, cells and viruses
Female 5-to 6 week old BALB/c (H-2d) mice were purchased from Samtako (O-San, Korea), and were maintained under standard conditions at the animal facility of Chonbuk National University, according to the Institutional Guidelines. All experiments were conducted according to the guidelines of the committee on the Care of Laboratory Animal Resources. HSV-1 McKrae strain was propagated in Vero cells (CCL81, ATCC, Manassas, VA) using DMEM supplemented with 2.5% FBS, penicillin (100 U/ml) and streptomycin (100 U/ml). The Vero cells were infected with HSV-1 at a multiplicity of infection (MOI) of 0.01, and were incubated in a CO2 incubator for 1 h at 37℃ with shaking at 15 min intervals. After absorption, the inoculum was removed, and 5 ml of maintenance medium containing 2% was added. Approximately 2~3 days post-infection, cultures of the host cells showing an 80~90% cytopathic effect were harvested, and were titrated by conventional plaque assay using Vero cells. The virus stocks were stored in aliquots at -80℃ until use.
Vaginal challenge
The mice were pre-treated with progesterone to synchronize their estrous cycles, as previously described (34). Briefly, mice were subcutaneously injected with Depo-provera (DP) at 2 mg per mouse. Five days following the injection of DP, the mice were challenged intravaginally with 106 pfu of HSV-1 McKrae strain per mouse, followed by intraperitoneal treatment of the indicated amino acid solution daily. The mice were examined daily for vaginal inflammation, neurological illness, and death, as described previously (34). Mice were scored 1 to 5 depending on the clinical severity of disease (0, no change; 1, mild inflammation; 2, moderate swelling; 3, severe inflammation; 4, paralysis; 5, death).
Virus titration in vaginal tract
Mice were injected sub-cutaneously with 2 mg of progesterone/mouse (Depo-Provera) to synchronize their estrous cycles, as described earlier (34). Five days following the injection of DP, the mice were challenged intravaginally with 106 PFU of HSV-1 McKrae. Vaginal washes were collected at days 2, 3 and 4, by pipetting 100 µl of PBS into and out of the vagina four to five times. Virus titers in vaginal washes were determined by plaque assay on a monolayer of Vero cells.
ELISA for IFN-γ expression in vaginal tract
The collected vaginal washes were used for determination of IFN-γ levels with ELISA. Briefly, ELISA plates were coated overnight at 4℃ with an optimal dilution (50 ng/well) of IFN-γ anti-mouse antibody (R4-6A2). The plates were washed three times with 0.05% PBS-Tween 20 (PBST) and were blocked with 3% skimmed milk. The samples were then serially diluted with twice their volume of PBS, and were incubated for 2 h at 37℃, alongside IFN-γ protein (Pharmingen, San Diego, CA) which was used as the standards. Following the addition of biotin conjugated anti-IFN-γ antibody, the plates were again incubated overnight at 4℃, and were subsequently washed and incubated at 37℃ with peroxidase-conjugated Streptavidin (Pharmingen) for 1 h. Color was developed by the addition of a suitable substrate (11 mg of 2, 2-azinobis-3-ethylbenzothiazoline-6-sulfonic acid in 25 ml of 0.1 M citric acid, 25 ml of 0.1 M sodium phosphate, and 10 µl of hydrogen peroxide). The concentration of IFN-γ was determined using an automated ELISA reader, and the SOFT max Pro4.3 program to compare with samples of standard cytokine.
Reagents and antibodies
L-Gln (G-3126), L-Leu (L8912), L-glutamic acid (Glu) (G8415), L-phenylalanine (Phe) (P5482), L-lysine (Lys) (L5501), L-asparagine (Asn) (A0884), and L-Arg were purchased from Sigma. All amino acids were dissolved in PBS (40mg/ml), and a volume of 200ul/mouse was administered twice daily via intra-peritoneal injection after the initial (Day 0) administration of HSV-1. The mAbs used for the flow cytometric analysis and other experiments were prepared from eBioscience (San Diego, CA) or BD Bioscience (San Diego, CA), and included: FITC-anti-mouse CD3e (145-2C11), CD4 (Rm4-5), CD8 (53-6.7) CD44 (IM7), CD62L (MEL-14), CD69 (H1.2F3), CD80 (16-10A1), phycoerythrin (PE) conjugated anti-mouse- CD4 (RM4-5) or CD8 (53-6.7), CD19 (MB19-1), CD154-(CD40L) (MR1), IFN-γ (XMG1.2), granzyme B (16G6), peridinin chlorophyll protein complex (PerCP)-conjugated anti-mouse IFN-γ, allophycocyanin (APC)-conjugated anti-mouse TNF-α, biotin-conjugated anti-mouse pan-NK cells (CD49b) [DX5], Streptavidin-APC.
Analysis of NK cell activity
Mice treated with Gln and Leu were sacrificed at 2 days post-infection, and splenocytes, iliac LN and vaginal cells were prepared. Prepared cells were stimulated with PMA and ionomycin in the presence of monensine (2 µM) to induce the expression of IFN-γ and granzyme B (PMA 50 ng/ml and ionomycin 150 ng/ml for 6 h). After being harvested, cells were treated with with FITC-CD3e, biotin-conjugated anti-mouse pan-NK cells (CD49b) [DX5] antibodies, and Streptavidin-APC for 30 min 4℃. Cells were then washed twice with FACS buffer containing monensin. After fixation, the cells were permeabilized with permeabilization buffer (eBioscience, San Diego, CA) and were stained intracellular with PE-anti IFN-γ (XMG1.2), and with granzyme B (16G6) in permeabilization buffer, each for 30 min at 4℃. Finally, the cells were washed twice with PBS and analysis was performed with a FACS Calibur flow cytometer.
Flow cytometric analysis for CD4/CD8 lymphocyte subsets
Groups of mice were challenged intravaginally with 106 PFU of HSV-1 McKrae and sacrificed at 5 days post-infection, after which splenocytes were prepared for flow cytometric analysis. The cells were stained by PE-CD4 or CD8, FITC-CD44, CD62L and CD69 antibodies. After washing, the cells were resuspended in PBS. Data collection and analysis were performed with a FACS Calibur flow cytometer (Becton Dickson Medical Systems, Sharon, MA).
Analysis of CD4+T cell responses specific to HSV-1
To monitor CD4+T cell responses specific to HSV-1, mice treated with Gln and Leu were sacrificed at 5 days post-infection and splenocytes, iliac LN and vaginal cells were prepared. The absolute number of IFN-γ, TNF-α producing CD4+ T cells specific to HSV-1 antigen was determined by the intracellular CD154 staining method, as previously described (35). In brief, freshly explanted splenocytes, iliac LN, and vaginal tract cells (2×106/well) were cultured in flat-bottom 96-well plates and were stimulated with UV-inactivated virus in the presence of PE-conjugated CD154 antibody for 12 h at 37℃. Monensin (2 µM) was added to the antigen-stimulated cells 6 h before harvest. Cell surfaces were stained for FITC-anti CD4 antibody for 30 min at 4℃ and were then washed twice with PBS containing monensin. After fixation, the cells were washed twice with permeabilization buffer (eBioscience, San Diego, CA) and were then stained intracellularly with PerCP-anti IFN-γ and APC-anti TNF-α in permeabilization buffer for 30 min at 4℃. Finally, the cells were washed twice with PBS and were analyzed with a FACS Calibur flow cytometer.
Statistical analysis
All analyses were performed using Graph Pad Prism software (Graph Pad Prism, San Diego, CA). Statistical analyses were performed using the Student's unpaired t-test. Also, Kaplan-Meier curves were analyzed by the log-rank test. Data represent the average±SD of levels derived from each group. p-values<0.05 were considered to be statically significant.
RESULTS
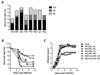
Glutamine and Leucine, but not their combinations, provide protection against mucosal infection of HSV-1
There is little information describing the supplement effect of amino acids against mucosal HSV-1 infection. To evaluate the protective efficacy of amino acids administration against a mucosal HSV-1 challenge, we selected several amino acids which included non-essential amino acid; Gln, Glu, Asn, and essential amino acid; Leu, Phe, Lys, Arg. Initially, we decided to examine the in vivo anti-herpes activity of all selected amino acids. To this end, we infected BALB/c mice with HSV-1 McKrae strain, and subsequently administered 4% solution of the above noted amino acid (200 ul/mouse) daily via intra-peritoneal route. The challenged mice were examined for the proportion of survival rate at 6, 9, and 12 days post-infection. Our results revealed that administration of Gln and Leu showed less mortality and significantly prolonged survival rates against mucosal challenge of HSV-1 at the indicated days, compared with other amino acids and control groups (Fig. 1A). To further understand the anti-herpes activity of Gln and Leu, we determined protective efficacy of 4% Gln and Leu administered once or twice daily, or as a combined mixture (200 ul/mouse) intraperitoneal injection after HSV-1 mucosal challenge. We observed that the intra-peritoneal administration of Gln(×2) and Leu(×1) resulted in the highest survival rate against mucosal challenge of HSV-1 (50~60%), and that surviving mice recovered rapidly and did not display any clinical severity. However, mice receiving their combined treatment (both ×1 and ×2) showed a moderately enhanced survival rate (20~30%), compared with control mice (10%) (Fig. 1B). Similarly, when the clinical severity was scored, Gln (×2) and Leu (×1)-treated mice showed significantly lower inflammation, but the group receiving their combined treatment showed identical inflammation to control mice (Fig. 1C). These results indicate that intraperitoneal administration of Gln (×2) and Leu (×1) could provide effective protection against mucosal HSV-1 infection, but that their combined treatment could not.
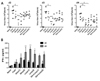
Gln and Leu reduce virus burden associated with increased levels of IFN-γ in vaginal tract following HSV-1 mucosal infection
To better understand the protective role of Gln and Leu treatment against mucosal HSV-1 infection, we examined the virus titers and production of IFN-γ in the vaginal tract. After mucosal HSV-1 infection, groups of mice were administered a daily 4% solution of Gln or Leu, or their combined mixture (all 200 µl) either once or twice daily, and the virus titer and IFN-γ levels in vaginal washes were compared with those of the control mice. Results indicated that intra-peritoneal administration of Gln (×2) and Leu (×1) induced a lower viral burden in vaginal washes, compared with other treated groups (Fig. 2A). Interestingly, we observed that the levels of shedding virus were significantly lower in groups administered Gln (×2) or Leu (×1) at 3 and 4 days post-infection, compared with control mice. Also, we determined the production of IFN-γ in vaginal lavages in response to HSV-1 infection (Fig. 2B). The Gln (×2)-treated group sowed a higher level of vaginal IFN-γ than the control, but Leu (×1)-treated mice were not different to the control in the production of IFN-γ. Interestingly, at 4 days post infection Gln (×2) and Leu (×1)-treated mice had significantly increased levels of IFN-γ determined in their vaginal washes 4 days after mucosal HSV-1 infection, compared to their combined treatment and control mice (p<0.01) (Fig. 2B). Also, our results revealed that treatment of 4% solution of Gln, Leu at optimal doses could reduce the virus burden associated with increased levels of IFN-γ in the vaginal tract following HSV-1 mucosal infection.
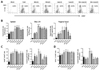
NK cell activity of Gln and Leu-treated mice following HSV-1 mucosal infection
NK cells have a major anti-viral effect through direct lysis of virus-infected cells and secretion of antiviral cytokines such as IFN-γ and TNF-α in the early stages of infection. Since Gln (×2) and Leu (×1) induced an increase in vaginal IFN-γ production, we checked whether NK cell activity was altered in Gln and Leu-treated mice after mucosal HSV-1 infection. Initially, we measured the frequency of CD3-DX5+ NK cells in the vaginal tract at 2 days post-infection. Our results indicated that NK cells are increased by intraperitoneal administration of Gln and Leu following HSV-1 infection. In particular, Gln(×2), Leu(×1), Leu (×2) and Gln+Leu (×1) groups increased NK cell populations in the vaginal tract, compared with other treated mice or control mice. In contrast, Gln+Leu (×2)-treated mice showed lower NK cell populations than control mice (Fig. 3A). Also, we determined the absolute number of NK cells in spleen, iliac LN and the vaginal tract at 2 days post-infection (Fig. 3B). The spleen of Gln (×2), Leu (×1), Leu (×2)-treated mice and iliac LN of Leu (×1), Leu (×2), Gln+Leu (×1), and Gln+Leu (×2)-treated groups had a higher absolute number of NK cells compared with control mice (p<0.01). In particular, the Gln (×2)-treated group showed significantly higher absolute NK cell numbers in iliac LN compared to control mice (p<0.001). We also found that the Leu (×2)-treated group had a higher absolute number of NK cells in the vaginal tract compared to control mice (p<0.01). The Gln (×2) and Leu (×1)-treated group showed a higher absolute number of NK cells in the vaginal tract, compared to control (p<0.05), but less than those of the Leu (×2)-treated group (Fig. 3B). To further understand the effect of Gln and Leu on NK cell activity, we checked the proportion and absolute number of granzyme B and IFN-γ-producing NK cells in spleen (Fig. 3C and D). The proportion of granzyme B-producing NK cells in Gln (×2), Leu (×1)-treated mice was significantly higher than those of control mice (p<0.05). Similarly, a significantly enhanced absolute numbers of granzyme B-producing NK cells in Gln (×2) and Leu (×1)-treated mice were observed. Consistent with this, the proportion and absolute number of IFN-γ-producing NK cells were observed to be significantly higher in Gln (×2), Gln (×1), and Leu (×1)-treated mice, compared to control mice. Therefore, these results indicate that the intra-peritoneal administration of Gln and Leu following HSV-1 mucosal infection could affect NK cell activity, thereby conferring efficient protective immunity.
Modulation of immune responses against HSV-1 mucosal infection by Gln and Leu
Beside NK cell activity, CD4+ T cell-mediated immunity has been known to play a crucial role in conferring efficient protective immunity against mucosal HSV-1 infection (34). To address whether Gln and Leu administration following HSV-1 infection could modulate a CD4+ T cell-mediated immune response in mucosal HSV-1, we analyzed the activation levels of splenic CD4+ and CD8+ T cells in response to HSV-1 infection (Table I). The intraperitoneal administration of Gln and Leu following HSV-1 mucosal infection showed more activated phenotypes of CD4+ and CD8+ T cells in the spleen. Also, we evaluated CD4+ T cell responses specific to HSV-1 antigen using intracellular CD154 staining. We harvested splenocytes, iliac LN, and vaginal tract cells from the control and all treated mice 5 days post infection, subsequently restimulating cells with UV-inactivated HSV-1 antigen in the presence of PE-conjugated CD154 antibody. After the stimulation, we evaluated the responses of CD4+ T cells by intracellular staining for IFN-γ and TNF-α. Our results revealed that the absolute number of IFN-γ+CD4+ T cells in the spleen of Leu (×2) and in iliac LN of Gln (×2), Leu (×2) and Gln+Leu (×1)-treated mice was significantly enhanced (Fig. 4A and B). Also, we found that the absolute number of TNF-α+ CD4+ T cells in spleen of Gln (both ×1 and ×2) and in iliac LN of Leu (×2) and Gln+Leu (×1)-treated group was significantly increased compared to that of the control group (p<0.01). Similarly, we found that the Leu-treated group showed a higher number of IFN-γ+TNF-α+CD4+ T cells in spleen and iliac LN. Interestingly, we recognized that the vaginal tracts of mice which received Gln (×2), showed higher absolute numbers of IFN-γ+, TNF-α+, and IFN-γ+TNF-α+CD4+ T cells, compared with other treated mice and control mice (Fig. 4C). Collectively, these results suggest that the intraperitoneal administration of Gln and Leu affect the protective immunity mediated by CD4+ T cells in responses to mucosal HSV-1 infection.
DISCUSSION
Several studies have shown that some amino acids function as building block of protein as well as regulators of maintenance, growth, reproduction and immunity. Here, we demonstrate that Gln and Leu have a regulatory function to provide protective immunity against mucosal infection of HSV-1. Our results indicate that intraperitoneal treatment of Gln and Leu is beneficial to induce protective immunity against HSV-1 mucosal infection, thereby conferring better survival rates upon treatment. Protective functions of Gln and Leu were further demonstrated by evaluating NK cell activity and HSV-1 antigen-specific CD4+ T cell responses in treated mice. Unexpectedly, the combined treatment of Gln and Leu did not provide better protective immunity than the treatment of individual amino acid. This implies that the treatment of Gln and Leu with overdose or combined treatment may be detrimental to the protective immunity against infectious disease which is conferred at lower single doses.
Several stuidies have demonstrated that Gln might play an important role in immunological function. Gln was known to influence Th1/Th2 responses (26). An optimal Gln concentration may be able to enhance the Th1 cytokine response, but little affect the Th2 cytokines response (26). In addition, our results can be supported by several further lines of evidence (23,26). It has been reported that Gln increased Th1 cytokine production (IL-2 and IFN-γ) in PHA-stimulated lymphocytes, and also that Gln enhanced Th1 cytokines (IFN-γ) levels, but not Th2 cytokine (IL-10) responses in BCG- or measles virus-stimulated lymphocytes. Similarly, our results suggest that a twice-daily administration of Gln and once daily administration of Leu to HSV-1-challenged mice showed enhanced NK cell activity and an increased number of IFN-γ and TNF-α-producing CD4+ T cells specific to HSV-1 antigen, which was associated with better survival. In particular, the twice daily administration of Gln and once daily administration of Leu showed a higher proportion of granzyme B and IFN-γ-producing NK cells following mucosal infection with HSV-1. Because Gln has also been found to be crucial for lymphoblastic transformation, plasma cell differentiation, and immunoglobulin synthesis by pokeweed mitogen stimulation (23), Gln administration could affect the proliferation of NK cells and lymphocytes in response to mucosal infection with HSV-1. The immunological function of Leu has been not established in detail. Here we found that Leu administration showed highly enhanced activity of NK cells which could affect protective immunity against mucosal infection of HSV-1. Although Gln and Leu is essential in the culture medium, it seems that higher Gln and Leu concentrations may have an inhibitory effect of the proliferative responses. Previous studies have shown that Gln at higher concentrations (more than 1 mM) inhibits lymphocyte proliferation stimulated by PHA, concanavaline A, pokeweed mitogen, or IL-2 (30,36). The possible explanation of the inhibitory effect by higher doses or frequencies of Gln and Leu, and their combined solution, could be due to the formation of Gln and Leu toxic metabolites, such as ammonia or glutamate, which contribute to the immunosuppressive effect (36). Therefore, it is assumed that the detrimental effect of higher frequent administration of Gln and Leu, and/or their combined administration, on the modulation of protective immunity may be induced by such speculations.
It is believed that NK cell activity and Th1-type CD4+ T cell responses play crucial roles in conferring efficient protective immunity against mucosal infections of HSV-1 (34). The source of the initial production peak of IFN-γ at 2 days post-infection is suspected to be NK cells rather than Th1-type CD4+ T cells, which are known to cause a later peak (34). We observed the levels of IFN-γ in the vaginal tract at 2 and 4 days after HSV-1 infection. As expected, the twice daily administration of Gln and once daily administration of Leu showed a potent protective immunity, inducing higher production of vaginal IFN-γ, compared to other treated groups (Fig. 2B). In parallel, the production of vaginal IFN-γ by treatment of Gln and Leu following HSV-1 mucosal infection was closely associated with viral burden in vaginal washes, which indicated that IFN-γ produced by NK and Th1-type CD4+ T cells was recruited into inflammatory tissues, and may be critical to control the outcome of diseases caused by HSV-1 mucosal infection. Interestingly, Gln administration induced higher numbers of IFN-γ and TNF-α-producing CD154+CD4+ T cells specific to HSV-1 antigen when enumerated antigen-specific CD4+ T cells recruited into vaginal tract. These results suggest that intraperitoneal administration of Gln and Leu following mucosal infection of HSV-1 initially increased NK-cell activity and subsequently Th1-type CD4+ T-cell activity, both of which were likely main sources of the soluble IFN-γ recovered in vaginal washes.
Although the issue of which immune defenses are involved in protection against disease following HSV-1 mucosal infection has not been fully resolved, the hypothesis of NK and Th1-type CD4+ T cells as the principal mediators is supported by several previous studies (34). Others, however, advocate that CD8+ T cells act as principal mediators for mucosal defense against HSV-1 infection (37). The present study favors the immunity correlated best with NK and Th1-type CD4+ T cell responses for providing efficient protective immunity against mucosal infection of HSV-1, even though the CD8+ T cell responses specific to the HSV-1 antigen were not addressed. In conclusion, our results suggest that intra-peritoneal administration of Gln and Leu provide a beneficial effect to modulate protective immunity against mucosal infection of HSV-1. However, dosage and frequency of administration should be carefully considered by the properties of infectious agents, because higher frequencies and overdose of Gln and Leu or their combined treatment showed detrimental effect sto protective immunity.


 XML Download
XML Download