

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abbreviations
APC
antigen presenting cell
BALT
bronchus-associated lymphoid tissue
CT-B
cholera toxin B subunit
DC
dendritic cell
Ig
immunoglobulin
GALT
gut-associated lymphoid tissue
GP2
glycoprotein-2
LP
lamina propria
MALT
mucosa-associated lymphoid tissue
MLN
mesenteric lymph node
MMP
matrix metalloproteinase
NALT
nasopharynx-associated lymphoid tissue
pIgR
polymeric Ig receptor
PP
Peyer's patch
PPL
Peyer's patch lymphocyte
SIgA
secretory IgA
TLR
Toll-like receptor
Treg
regulatory T
UEA-1
Ulex europaeus agglutinin 1
INTRODUCTION
Vaccination has played a central role in decreasing child mortality and increasing life expectancy (1,2). Vaccination has also resulted in the complete eradication of smallpox and a dramatic reduction in diseases caused by several infectious agents including bacteria and viruses. Most current vaccines are parenteral vaccines that are injected, although the principle of oral mucosal vaccination was first introduced with the Sabin oral polio vaccine that was developed in the early 1960s (3). Oral immunization is considered an ideal strategy for vaccination due to advantages such as needle-free application of the vaccine and ability to induce protective immunity in the mucosal area, a frontline of pathogen infection, which is not feasible by parenteral immunization (4). A representative licensed oral mucosal cholera vaccine is Dukoral®, which comprises killed whole Vibrio cholera and the recombinant cholera toxin B subunit (CT-B). This vaccine is effective at inducing immunity as shown by the induction of secretory IgA (SIgA) against both V. cholera lipopolysaccharide and CT-B. It also triggers herd immunity capable of reducing the spread of infection of the bacteria and induces long-term B- and T-cell memory in both mucosal and systemic immune compartments (5,6).
Although the usefulness of oral mucosal vaccines is not disputed, only a handful of licensed mucosal vaccines are commercially available. This is because inefficient antigen delivery across epithelial cells into mucosa-associated lymphoid tissue (MALT) and the tendency of the oral mucosal immune system to induce tolerance rather than immune stimulation act as intrinsic barriers to the induction of effective immune responses (7,8). In the late 20th century, mucosal immunology was a niche discipline due to limited understanding of the mucosal immune system. However, the constant efforts of immunologists over the last three decades have strengthened our understanding of the working and regulation of mucosal immune responses, and mucosal immunology has evolved to be an important area of immunology and vaccinology (9). Consequently, we are now in good position to understand the mucosal immune system and to develop effective mucosal vaccines. In this review, we summarize current knowledge of the mucosal immune system and mucosal vaccine development, with an emphasis on mucosal vaccine adjuvants.
MUCOSAL IMMUNE SYSTEM
Mucosal surface and mucosal epithelial cells
Mucosal surfaces cover ~400 m2 of the human body surface and function both as physical barriers and in immune defense against pathogen invasion (10). The mucosal area maintains its integrity through coordinated interactions among the microbial flora, the physical barrier properties of the mucosa, and immune defense. Mucosa are directly exposed to the external environment and many pathogens utilize the mucosal surface to invade the body when its integrity and balance are disrupted (11,12). Mucosal surfaces are classified into two types based on their distinct characteristics. Type I mucosal surfaces are found in the gut alimentary, respiratory, and female upper reproductive systems, while type II mucosal surfaces are found in the visual, mouth alimentary, and female lower reproductive systems (13). Type I mucosal surfaces have a simple columnar epithelial layer in which IgA is the major immunoglobulin (Ig) isotype. In contrast, type II mucosal surfaces have a stratified squamous epithelial layer, and the major Ig isotype is IgG.
The gastrointestinal tract, which is lined by type I mucosa, is the best studied tissue among the diverse mucosal systems with respect to the variety of cells in the mucosal immune system. Gut epithelia contain all five epithelial lineage cells: goblet cells, Paneth cells, M cells, enterocytes, and intraepithelial immune cells. Goblet cells are specialized epithelial cells that secrete mucin glycoproteins that separate the gut microbiota from epithelial cells, and also provide anchorage sites for and nutrients to microbiota (14). The function of goblet cells is determined by the characteristics of the mucins they secrete. Mucins form a viscous gel-like bilayer that extends up to 150µm from the intestinal epithelial cell surface. The firm inner layer comprises protective components against microbes, while the loose outer layer contains huge numbers of bacteria (15). The inner layer impedes bacterial penetration due to the presence of protective components such as IgA and antimicrobial proteins, including defensins and C-type lectins. The critical role of mucus as an immune regulator was demonstrated in a mouse model with a missense mutation in the Muc2 gene (Winnie mouse). Mice with this mutation developed chronic inflammation in their distal intestines resembling human ulcerative colitis (16). In addition, the finding that Muc2 gene expression was modulated by short-chain fatty acids, which are the end-products of microbial fermentation, suggested a close relationship between the microbiota and host mucosal epithelia. Similarly, when Dicer, an enzyme involved in miRNA maturation, was knocked down only in intestinal epithelial cells, goblet cell differentiation was defective, and the host became more susceptible to parasite infection (17).
Paneth cells are specialized secretory epithelial cells located in the epithelium of the small intestine that have large secretory granules containing a wide variety of proteins (18). These cells are continuously renewed from a common epithelial stem cell population in the crypts and are regulated by an unfolded protein response activated by endoplasmic reticulum stress originating from autophagy (19,20). Paneth cells secrete antimicrobial peptides (α-defensins and cryptdin-related sequence peptides), antimicrobial C-type lectins (REGIIIα), β-1,4-glycosidases (lysozyme C), and phospholipid-sn-2 esterases (sPLA2) that are effective at inhibiting the growth of Gram-positive and/or -negative bacteria. The function of Paneth cells is closely associated with intestinal diseases such that the number of translocated commensal bacteria increases when Paneth cells are removed by the cell-specific expression of diphtheria toxin (18). The next cell type is enterocytes, which function mainly as an epithelial barrier. These cells also inhibit Th1 cell differentiation and modulate tolerogenic CD103+ dendritic cells (DCs) through the secretion of TGF-β, retinoic acid, and thymic stromal lymphopoietin (21,22). In this process, BAFF and APRIL produced by enterocytes promote IgA class-switching in B cells (23).
M cells are specialized epithelial cells capable of performing transcytosis of luminal antigens. M cells are located in the follicle-associated epithelium of Peyer's patches (PPs) and nasopharynx-associated lymphoid tissue (NALT), as well as in isolated lymphoid follicles and the appendix (24). Recently, the presence of an inducible M cell population at the tips of intestinal villi was reported - so-called villous M cells (25). M cells were first identified in 1965 from the rabbit appendix by Schmedtje and were named 'M' cells by Owen and Jones because of the presence of 'microfolds' on their apical surface (26,27). The origin of M cells has been subject to controversy, but it is accepted that both enterocytes and M cells originate from a common precursor, crypt cells, which are stem cells located between the villus and the dome of PPs (28). However, it is not clear whether crypt cells differentiate directly into enterocytes and M cells or whether other factors act later to induce further differentiation of pre-formed enterocytes into M cells. The former possibility is supported by the finding of M cell precursor cells in specialized dome-associated crypts of PPs (29). The latter possibility is supported by the finding that functional M cell development, at least in the intestine, requires several factors such as the presence of B lymphocytes or the expression of CD137 or RANK ligand (30-32). M cells play a critical role as gatekeepers in the mucosal immune system, although only 1 of every 10 million epithelial cells in the intestinal tract is an M cell (33). In fact, although the mucosal epithelium consists of a single layer, the thick glycocalyx layer reduces direct contact between epithelial cells and microbes. However, the surface of M cells not only has a very thin glycocalyx layer, but also short and irregular microvilli (34). In addition, M cells contain docking sites for lymphocytes and antigen presenting cells (APCs), which is important for stimulating specific mucosal immune system. These features of M cells make them attractive targets for pathogen invasion and vaccine delivery (35).
Although it is well-known that M cell-mediated transcytosis of luminal antigens is very efficient and rapid, the specific mechanisms of antigen processing and presentation in M cells are not yet completely understood. For example, it is unclear whether M cell-mediated antigen uptake is a specific receptor-mediated process or not (26). In addition, the observation that M cells have a 16-fold decreased lysosomal volume and low lysozyme activity compared to normal enterocytes have led researchers to question whether M cells have a direct role in antigen processing. However, it is believed that M cells may possess some enzymatic activity, such as cathepsin E activity, which is normally found in the lysosomal compartment of APCs (36). Our limited understanding of the biology of M cells is due to the challenges associated with identification and isolation of M cells, because M cells comprise only approximately 5% and 10% of the intestinal lymphoid follicle surface area in humans and mice, respectively. Fortunately, an in vitro M-like cell culture model comprising Caco-2, a human adenocarcinoma cell line, and Peyer's patch lymphocytes (PPLs), suggested by Kernéis et al. in 1997 and further modified in 2000 by using the human B cell line Raji instead of PPL, has facilitated investigations into M cells (37,38). This in vitro M-like cell model accelerated the study on antigen transport across M cells, identification of M cell-specific molecules, and elucidation of interactions between M cells and bacterial and vaccine materials.
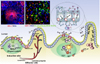
Structural features of the mucosal lymphoid organ
The mucosal immune compartment comprises two distinct structures: inductive MALT and effector sites (Fig. 1). MALT comprises solitary organized mucosa-associated lymphoid follicles that contain B cell follicles, an inter-follicular T cell area, and a variety of APCs, and does not have afferent lymphatics. MALT includes gut-associated lymphoid tissue (GALT) comprising PPs, the appendix, and isolated lymphoid follicles (ILFs), nasopharynx-associated lymphoid tissue (NALT) that comprises adenoids and the paired palatine tonsils, and bronchus-associated lymphoid tissue (BALT). Although the mesenteric lymph node (MLN) in the small intestine is associated with mucosal immune induction, it is different from GALT in its structure, because MLNs are connected with afferent lymphatics (39). Mucosal immune effector sites comprise the lamina propria (LP) and surface epithelium of the gut, although the effector sites are not always clearly distinguishable from inductive sites (39).
Secretory IgA (SIgA) in mucosal immunity
Serum IgA was discovered by Grabar and Williams in the early 1950s, and it was later shown that IgA is the dominant immunoglobulin isotype in mucosal secretion (40). In particular, a report that SIgA recognizes antigenic epitopes different from those recognized by serum Ig made SIgA an attractive target for vaccination outcome against many pathogens (41). Mucosal SIgA is produced via two distinct pathways, namely T cell-dependent and -independent pathways (42). T cell-dependent induction of IgA+ B cell differentiation occurs in the germinal center of PPs; DCs located near M cells encounter antigen and migrate to the T cell zone and activate T cells (Fig. 1). Activated follicular T cells express CXCR5 and migrate toward CXCL13 in the follicular DC network where activated T cells help activate B cells. B-T cell interactions through MHC-TCR and CD40-CD40L are critical for B cell activation, induction of activation-induced cytidine deaminase, and class switch recombination/somatic hypermutation in the germinal center of PPs. After up-regulation of gut-homing receptors, such as CCR9, α4β7 integrin, and type 1-sphingosine 1-phosphate, IgA+ plasmablasts migrate from PPs to the gut LP where they further differentiate into IgA-secreting plasma cells that secrete dimeric IgA. Dimeric IgA is transported into the gut lumen by the interaction of the secretory component of dimeric IgA with polymeric Ig receptors (pIgR) expressed on the basolateral surface of epithelial cells (43). In contrast, T cell-independent expression of IgA occurs mainly in ILFs and the LP. iNOS+ DCs stimulated by luminal bacteria express TNFα, which induces the expression of matrix metalloproteinases (MMPs). MMPs such as MMP9 and APRIL convert latent TGF-β1 into its active form, and active TGF-β1 facilitates preferential class switching from IgM to IgA in B cells. In fact, there is a large amount of BAFF, APRIL, IL-6, and IL-10 present in the LP environment, which facilitates the survival, proliferation, and differentiation of IgA+ B cells into plasma cells. SIgA in the mucosal compartment has several biological functions including maintenance of homeostasis of the mucosal system in response to pathogen infection (44). For example, immune exclusion by SIgA can limit interactions between the epithelium and microoragnisms, thereby blocking host invasion by pathogens. In addition, SIgA has antimicrobial effects; dimeric SIgA is effective at microbial agglutination and virus neutralization (43).
Lymphoid cellular components in mucosal immunity
Although SIgA, Th1, Th2, and Th17 cells are primarily responsible for mucosal immune responses, cytotoxic CD8+ T cells are also important in the mucosal immune response. In vaccine-induced immune responses, protection against various bacterial and viral infections depends on Th1 cell activation, whereas stimulation of Th2 cells is associated with antibody production (45). Th17 cells, which are located mainly in the mucosal LP, are implicated in protection against mucosal pathogens and play a role in memory cell induction (46,47). Finally, cytotoxic CD8+ T cells produced in response to vaccination show cell-mediated immunity, although their activity is not yet fully understood (48).
MUCOSAL VACCINATION
Mucosal vaccine development
Mucosal vaccines, especially oral mucosal vaccines, have several advantages over systemic vaccines such as ease of administration without the need for a needle or trained personnel. These features of oral mucosal vaccines make them attractive for mass immunization during pandemics (49). However, licensed mucosally administrable vaccines are available only for cholera, Type A and B influenza viruses, poliovirus, rotavirus, and Salmonella Typhi (9). One of the most significant problems associated with oral mucosal vaccines is possible tolerance induction against the orally introduced antigens, because it is generally known that oral administration of soluble proteins dominantly triggers oral tolerance (50,51). Tolerance induction by orally-introduced vaccine materials translates into systemic unresponsiveness after challenge infection by the same antigen. To overcome this tolerogenic mucosal environment, many efforts have been made to develop effective mucosal adjuvants that can stimulate both innate and adaptive immunities and are capable of inducing effective mucosal and systemic immune responses. The next hurdle in mucosal vaccine development is the unique compartmentalization and preferential dissemination of immune responses after vaccination through each route (52). Irrespective of the antigen sampling mechanism, mucosally-introduced antigens trigger B and T cell activation in inductive sites such as NALT and PPs, and then antigen-activated lymphocytes home to selected sites according to homing and chemokine receptors including α4β7, CCR9, and CCR10 (53,54). Therefore, specific mucosal compartmentalization determines the dissemination of established mucosal immune responses. For instance, oral immunization may induce an antigen-specific immune response in the proximal part of the small intestine, ascending colon, stomach, and in mammary and salivary glands (52). Consequently, a holistic approach is required to construct effective immunity by ensuring effective delivery of vaccine materials to proper mucosal inductive sites and by linking the innate and adaptive immune response by blocking oral tolerance induction.
Mucosal vaccine adjuvants
Vaccine adjuvants, especially in mucosal vaccines, can support the induction of antigen-specific immune responses in a tolerogenic microenvironment. In addition, adjuvants are closely associated with the induction of high-affinity and/or long-lived memory B cell differentiation (9). In fact, adjuvant activity results from bridging the divide between innate and adaptive immunity and, consequently, ligands for pattern recognition receptors such as Toll-like receptors (TLRs) are considered primarily as mucosal adjuvants (55,56). For example, the commercial IC31 adjuvant contains a synthetic oligodeoxynucleotide (ODN1a), a TLR-9 agonist, and is able to induce both Th1- and cytotoxic T cell-mediated immunity (9). Monophosphoryl lipid A, muramyl dipeptide, and flagellin, which are ligands for TLR-2, -4, or -5, respectively, are also used as adjuvants in mucosal vaccines (56). Enterotoxin is another important component of some mucosal adjuvants. For example, enterotoxins such as cholera toxin and E. coli heat-labile toxin are expected to enhance immune responses by activating the innate immune system. Moreover, they bind to gangliosides via their B subunits and enhance T cell-mediated immune responses by stimulating DCs. Chitosan, Quillaja saponins, and cytokines such as IL-1 and IL-12 are other mucosal adjuvant candidates.
Mucosal vaccine delivery systems
As explained above, securing antigen delivery to the proper mucosal immune inductive site is important for developing a successful mucosal vaccine. Mucosal vaccination was initially performed through the oral route. Currently, however, investigators are exploring intranasal, aerosol inhalation, intravaginal, rectal, and sublingual routes for mucosal vaccination, although only oral and intranasal routes have been licensed for vaccine applications in humans (55). Oral mucosal vaccines are directly exposed to the harsh gut environment and an immune tolerogenic environment (56). To address these challenges, several strategies have been developed. For example, live attenuated bacteria such as Salmonellae, E. coli, Lactobacilli, Shigella, Listeria, Mycobacteria, and Streptococci and viruses such as poxviruses, adenoviruses, polioviruses, herpesviruses, mengoviruses, and influenza have been considered as delivery vehicles for mucosal vaccination. These attenuated pathogens can serve the dual functions of delivering antigen and inducing effective adaptive immunity via their strong, intrinsic adjuvant activity (55,57). The representative licensed vaccines that use this strategy are the rotavirus vaccine and oral Ty21A typhoid vaccine. However, the stability and safety of live attenuated microbe vaccines have been questioned. In that sense, antigen-expressing plant-based edible vaccines are an interesting alternative oral mucosal vaccine delivery system because of the ability to perform cold-chain- and needle-free vaccination (58-60). In particular, given that transgenic plant-based vaccines can now be produced cost-effectively and do not require cold preservation or professional healthcare workers for administration, they are considered ideal vaccination systems in third world and developing countries. For example, when experimental MucoRice was administered orally, specific immune responses against cholera toxin in both systemic and mucosal areas were detected (61). Finally, particles such as liposomes, dendrimers, multiple emulsions, immune stimulating complexes (ISCOMs), and biodegradable polymers have been added to vaccines to enhance the efficiency of vaccine delivery (62,63). For example, oral administration of liposomes containing influenza antigen enhanced the Th1-biased response, serum IgG2a production, and antigen-specific IFN-γ production (64). This strategy for vaccine delivery was shown to have specific targeting ability through association with M cell-targeting ligands (65).
M cell-specific targeting ligands
Understanding the characteristics of M cells could provide crucial information for efficient oral mucosal vaccine development, because M cells are the main gateway not only for mucosal antigens, but also for various pathogens such as Salmonella spp., Yersinia enterocolitica, Shigella spp., Vibrio cholera, reovirus type-1, poliovirus, and HIV-1 (65). Previously, when little was known about M cell markers, M cells were distingushied based only on morphological characteristics such as a thin glycocalyx layer, short and irregular microvilli, and an inner pocket (66). Recent intensive endeavors to determine the molecules expressed uniquely by M cells have resulted in the identification of targets that can be exploited for mucosal vaccine delivery (Table I). Both Ulex europaeus agglutinin 1 (UEA-1) lectin and Aleuria auranitia bind specifically to α1,2-fucose residues expressed on the apical surface of mouse M cells (67,68). These interactions between lectin and the surface molecules of M cells allow the selective binding and rapid uptake of conjugated antigens into M cells. For example, when PLGA nanoparticles or liposomes containing both UEA-1 and hepatitis B surface antigen were administered orally, they predominantly targeted PP M cells and elicited significantly higher SIgA responses than particles containing only UEA-1 (69,70). However, the drawbacks of using plant lectins in oral vaccines include their toxicity and possibility of degradation once anchored on the surfaces of microparticles (71). In a later study, mouse M cells isolated using UEA-1 were used to develop a M cell-specific antibody, NKM 16-2-4, which itself results in an efficient delivery of antigen into M cells (72). Similarly, galectin-9, which binds to N-glycans and repeated oligoactosamines on human M cells, can enhance antigen delivery, although galectin-9 induced Treg-skewed immune responses (24,73). Collectively, the results from studies that investigated M cell-specific lectins and antibodies against M cell-specific receptors suggest that the M cell-targeting of antigens could be used for creating effective mucosal vaccine delivery systems.
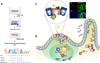
The lessons from the study for identifying M cell-targeting ligands include that simple targeting of antigen to M cells does not guarantee the induction of efficient protective immunity. Consequently, pathogen recognition receptors, such as TLR-4 and platelet-activating factor receptor that are involved in the onset of innate immune, have been considered as targeting receptors for oral mucosal vaccines (74,75). In addition, a thorough assessment of the immune responses induced by an M cell-targeting strategy is essential to determine the efficacy of the M cell-targeting strategy. Our research group has attempted to select M cell-targeting ligands with both M cell-targeting and adjuvant activities by using a screening phage display library against an in vitro M-like cell culture model (Fig. 2) (76). When the M cell-targeting ligand Co1 (SFHQLPARSPLP) was used in an oral dengue vaccine, it not only effectively delivered the conjugate antigen, the envelope protein III of dengue virus, both in vivo and in vitro to the apical surface of murine M cells, but also evoked antigen-specific immune responses in both mucosal and systemic compartments. In addition, a Th2-skewed immune response and non-tolerogenic characteristics were induced. Consequently, M cell-targeting ligands might increase the uptake of orally administered vaccines by M cells and enhance antigen-specific immune responses in both mucosal and systemic compartments.
Discovery of M cell-targeting receptors using pathogen-exploited molecules
Efforts to identify the molecules exploited by pathogens to invade M cells have focused on identifying M cell-specific targeting molecules that can potentially be used in mucosal vaccines (35). For example, polystyrene nanoparticles coated by Yersinia-derived invasin showed enhanced antigen-targeting and antigen-specific immune responses through interaction with β1 integrin expressed on the apical surface of M cells (77). When σ1 of reovirus, which preferentially invades M cells, was applied in a mucosal DNA vaccine, it evoked mucosal immune responses through interaction with α2,3-sialic acid expressed on the apical surface of M cells (78). An additional study identified glycoprotein-2 (GP2) in human and mouse M cells through gene expression profiling as a receptor for FimH+ bacteria, indicating that GP2-specific antibodies could potentially be of use in mucosal delivery and adjuvant systems (79). Recently, it was reported that the cellular prion protein PrPc is expressed on M cells and that it is exploited by Brucella abortus (80). We also identified the expression of the complement C5a receptor (C5aR, CD88) in M cells while searching for the receptor for M cell-targeting ligand, Co1. The Co1 ligand has 60% amino acid sequence homology with the OmpH α1 helix of Y. enterocolitica, which has a similar sequence to skp of E. coli, which in turn is a ligand for C5aR (81). We showed that C5aR is expressed on the apical surface of both human M-like cells and mouse M cells and that it plays a non-redundant role during Y. enterocolitica infection or antigen uptake into M cells (81). These studies suggest that M cell-mediated antigen transcytosis could be described, at least in some part, as a receptor-mediated process (9,82). Together, these results indicate that knowledge of the receptors on the apical surface of M cells that are involved in pathogen invasion could be used to design M cell-targeting ligands with adjuvant activity.
CONCLUSIONS AND FUTURE PERSPECTIVES
Mucosa, which are exposed to heavy loads of commensal and pathogenic microorganisms, represent a first line of defense against pathogen invasion. Although the parenteral vaccines currently in use are effective at protecting against many pathogens, mucosal vaccines have several advantages over parenteral vaccines. However, immune tolerance to maintain homeostasis is a hallmark of the mucosal environment. Despite the very low number of M cells in the mucosal epithelium, the ability of M cells to sample antigens, translocate bacteria, and initiate mucosal immune responses make them attractive targets for mucosal vaccine delivery. Therefore, to develop mucosal vaccines, safe and effective mucosal adjuvants capable of M-cell targeting and with adjuvant activity are required. In this review, we summarized the current state of knowledge of the structure of the mucosal immune system and the M cell-targeting strategies that have been explored to develop effective oral mucosal vaccines. We anticipate that further understanding of mucosal immunology and M cell biology will allow the development of the next generation of mucosal vaccines.

 XML Download
XML Download