

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Recently, a great advance was achieved in the understanding of pathogenesis of autoimmune diabetes, particularly regarding the molecular mechanism of pancreatic β-cell apoptosis and immunological significance of β-cell apoptosis. Hence, it has been shown that pancreatic β-cell death is important not only in the final effector phase of autoimmune type 1 diabetes (1,2) but also in the initiation of β-cell autoimmunity (3,4). Such progress was mostly achieved by employing murine models of autoimmune diabetes because application of most essential genetic manipulation or immunological intervention is possible only in animal models. Murine autoimmune diabetes models and human type 1 diabetes have many common characteristics such as the presence of insulitis, requirement for specific MHC haplotypes and autoimmune responses to autoantigens including glutamic acid decarboxylase (GAD). Thus, many recent progress obtained employing murine autoimmune diabetes models would be applicable to human type 1 diabetes, implying important therapeutic potential. Autoreactive T lymphocytes are the most important effector cells in murine autoimmune diabetes (5-7), and probably in human type 1 diabetes. They will ultimately induce apoptosis of β-islet cells in murine autoimmune diabetes (8,9), while apoptosis of pancreatic β-cells in human type 1 diabetes has not been demonstrated because of the difficulty in procuring human pancreatic tissue and inability to synchronize disease process in human type 1 diabetes.
We have shown that IFNγ/TNFα synergism is responsible for murine pancreatic β-cell apoptosis both in vitro and in vivo (2), which was critically dependent on STAT1 activation by IFNγ (1). In vivo role of STAT1 activation by IFNγ was also demonstrated using STAT1-knockout NOD mouse model (1). Other death effector molecules such as Fas ligand (FasL) might be an effector molecule for a small number of autoreactive lymphocytes (10) but not for the majority of final death effector cells (9,11). IFNγ appears to sensitize otherwise resistant pancreatic islet cells or insulinoma cells to TNF α-mediated apoptosis by activating STAT1/IRF-1 signal pathway (2). Most such pancreatic β-cell death and signal pathways were studied in murine autoimmune diabetes models. Such findings might also be relevant in human type 1 diabetes also. In an effort to elucidate the possible mechanism of human pancreatic islet cell apoptosis leading to human type 1 diabetes, we investigated if similar cytokine synergism could induce apoptosis of human pancreatic islet cells and similar signal molecules are induced in them.
MATERIALS AND METHODS
Isolation of human islet cells
Human pancreatic islets were obtained from brain-dead patients between 1998 and 2002 at Samsung Medical Center, using a modification of the automated method for human islet isolation (12). Briefly, 350 ml of Hank's buffered salt solution (HBSS) containing 9.1µmol/l collagenase solution (Liberase, Boehringer-Mannheim, Mannheim, Germany) was injected through the pancreatic duct after cannulation. The pancreas was loaded into a stainless steel digestion chamber, and islets were separated during a continuous digestion process for 30~45 min. During the recirculation phase (flow rate, 85 ml/min), intrachamber temperature was increased at a rate of 2℃/min by the passage of the solution through a stainless steel coil immersed in a water bath (50℃). The chamber containing the distended pancreas was gently agitated and samples were taken every 2 min to monitor digestion. After 20~30 min of recirculation, digestion was stopped by dilution in 400 ml/min ice-cold HBSS. In this phase, the digested tissue was rapidly collected in sterile bottles containing 200 ml FCS. Purification of the islets was carried out over discontinuous Euro-Ficoll gradients (1.108, 1.096, 1.037 and HBSS). Purified islets were then washed twice, evaluated for purity, and counted after dithizone (Sigma, St. Louis, MO) staining. To make single islet cells, islets were spinned at 1,000 rpm for 2 min. The pellet was resuspended in 3 ml warm 53.7µmol/l trypsin-3 mmol/l EDTA. After incubation at 37℃ for 5 min and pipetting for 2~3 min, islets became invisible. Then, cells were washed with warm RPMI-10% FCS at 1,500 rpm for 5 min. Single islet cells were frozen in liquid nitrogen until use. At the time of assays, single islet cells were thawed and 1.4×105 cells were plated per single well of 96-well plates. Viability of the islet cells after thawing was above 90%, as judged by trypan blue staining and acridine orange/propidium iodide staining. Informed consent was obtained from the family members of the patients. Isolated human pancreatic islet cells were grown in DMEM containing 15% FBS, 2 mM glutamine, and penicillin-streptomycin (Gibco-BRL, Gaithersburg, MD). Recombinant human IFNγ was from Green Cross (Yongin, Korea). Recombinant human TNFα and recombinant human IL-1β were purchased from R&D Systems (Minneapolis, MN). All other chemicals were from Sigma Chemical Co., St. Louis, MO) unless stated otherwise.
Assessment of cytotoxicity by MTT assays
Cells (2×104/well for human single islet cells) were seeded in 96-well microtiter plates and treated with various combinations of cytokines for the indicated time periods. The optimal concentrations of cytokines for the cytotoxic action were 1,000 U/ml for IFNγ, 10 ng/ml for TNFα, and 17.5 ng/ml for IL-1β. After cytokine treatment, the medium was removed and 3-[4,5-Dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide (MTT, 0.5 mg/ml) was added followed by incubation at 37℃ for 2 hr in CO2 incubator. After a brief centrifugation, supernatant was carefully removed and DMSO was added. After insoluble crystals were completely dissolved, absorbance at 540 nm was measured using Thermomax microplate reader (Molecular Devices, Sunny Vale, CA).
Morphological analysis of apoptotic cells
Morphological changes in the nuclear chromatin of cells undergoing apoptosis were detected by staining with 2.5µg/ml of DNA-binding bisbenzimide Hoe33342 fluorochrome (Calbiochem, San Diego, CA), followed by an examination on a fluorescence microscope. For transmission electron microscopy, cells were fixed in 4% glutaraldehyde/1% paraformaldehyde/0.2 M phosphate (pH 7.2) at 4℃ for 2 hr. After two washes in 0.2 M phosphate, the cell pellets were postfixed with 2% OsO4 in the same buffer for 30 min. The pellets were dehydrated in ethanol and then in 100% propylene oxide, followed by embedding at 37℃ overnight and 60℃ for another 3 days. Ultrafine sections were cut and examined on an electron microscope (Hitachi H7100, 75 kV).
Western blot analyses
Cells were lysed in triple-detergent lysis buffer (50 mM Tris-Cl, pH 8.0, 150 mM NaCl, 0.02% sodium azide, 0.1% SDS, 1% NP-40, 0.5% sodium deoxycholate, 1 mM PMSF). Protein concentration in cell lysates was determined using Bio-Rad protein assay kit. An equal amount of protein for each sample was separated by 10% or 12% SDS-PAGE and transferred to Hybond ECL nitrocellulose membrane (GE Healthcare Life Sciences, Buckinghamshire, UK). After blocking with 5% skim milk, the membranes were sequentially incubated with one of the primary antibodies (rabbit anti-mouse IRF-1, Santa Cruz; rabbit anti-phospho-STAT1, New England Biolabs, Ipswich, MA) and then HRP-conjugated secondary antibodies (anti-rabbit IgG, GE Healthcare Life Sciences, Buckinghamshire, UK), followed by ECL detection (GE Healthcare Life Sciences, Buckinghamshire, UK).
RESULTS AND DISCUSSION
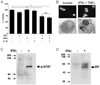
Treatment of single human islet cells with cytokines disclosed that combination of IFNγ, TNFα and IL-1β induced significant death after 5 days of incubation. When the effect of individual cytokine was studied, a combination of IFNγ and TNFα induced death of human islet cells while single cytokine did not exert significant cytotoxicity on human islet cells (Fig. 1A). The addition of IL-1β to the IFNγ/TNFα combination had only a minor effect on islet cell viability. Our results showing death of human islet cells by the triple combination of IFNγ, TNFα and IL-1β is similar to previous reports by others employing human islet cells (13-16). Percent death of human islet cells by the cytokine combination in this investigation was similar to a previous report (13), and was smaller than that of murine islet cells by the similar cytokine combination even after prolonged incubation for 5 days (2). Most of the death of human islet cells by the triple combination could be explained by IFNγ/TNFα combination because the addition of IL-1β to the IFNγ/TNFα combination increased human islet cell death only to a small degree, consistent with a previous paper (13).
Hoechst 33342 staining and electron microscopic examination showed that the death of human islet cells by IFNγ/TNFα combination was a typical apoptosis characterized by nuclear condensation and fragmentation with preserved plasma membrane integrity (Fig. 1B). These results are similar to previous reports showing nuclear condensation or TUNEL+ nuclei after cytokine treatment of human islet cells (15,16).
Next we asked if IFNγ activates STAT1 and induces IRF-1 which has been reported to be important for induction of TNFα susceptibility in otherwise resistant murine islet cells or cancer cells (2,17). Immunoblot analysis using antibody to phosphorylated STAT1 showed that STAT1 became activated 30 min after IFNγ treatment, similar to murine insulinoma cells (2) (Fig. 1C). IRF-1 was also induced by IFNγ treatment of human islet cells for 48 hrs again similar to murine insulinoma cells suggesting that STAT1 activation and IRF-1 induction by IFNγ may sensitize human islet cells to TNFα-induced apoptosis, similar to the case of murine islet cell death by IFNγ/TNFα synergism (2) (Fig. 1D). These results were similar to our previous study using murine islet/insulinoma cells and suggests the possibility that intracellular signal activated by IFNγ may sensitize otherwise resistant human pancreatic islet cells to TNFα-induced apoptosis, which could not be proved because of shortage of human islet cells, the absence of immortalized human islet cells and unavailability of genetic manipulation system. A recent paper has shown that Bim, a BH3-only proapoptotic Bcl-2 family member, is induced in human islets or rat islet cells after treatment with IFNγ+TNFα which plays an important role in β-cell death by IFNγ+TNFα synergism (18). In that model, Bim induction was attributed to IFNγ-induced activation of STAT1 which was bound to Bim promoter (18). While detailed biochemical and cellular mechanism of human islet cell death by cytokines could not be investigated further because of limited availability of human islets for diabetes research, our results suggest that the death of human islet cells is basically similar to that of murine islet cells.
 XML Download
XML Download