

 ePub
ePub Citation
Citation Print
Print


INTRODUCTION
CD4+Foxp3+ regulatory T cells (Tregs) make up 5~10% of peripheral CD4+ T cells and are essential for maintaining peripheral self-tolerance (1). The transcription factor Foxp3 is not only a unique marker for Tregs but also is required for the development and activity of the Tregs. The importance of Tregs in preventing autoimmunity has been proven in scurfy mice and in human patients with immunodysregulation, polyendocrinopathy, enteropathy, X-linked syndrome (IPEX) (2,3). Both the mice and human mutants lack functional Tregs due to loss-of-function mutations in the Foxp3 gene, and the absence of these cells is accompanied by severe autoimmune responses affecting many organs. In addition, quantitative and functional defects in Tregs are associated with autoimmune disorders, such as type 1 diabetes, multiple sclerosis and systemic lupus erythematosus (4-6).
Tregs elicit their regulatory effects on other immune cells by diverse mechanisms. They were originally identified by finding that they inhibited the proliferation of other T cells as well as cytokine production by the latter (7). They also regulate the maturation of dendritic cells (DCs), so preventing T cell priming (8). Their suppressive activity depends on cell-to-cell contact and/or the production of cytokines. Apart from secondary lymphoid tissues, Tregs are also found at sites of inflammation, such as arthritic synovial tissue and diabetic pancreases, although their roles such locations remain unclear (9,10).
Rheumatoid arthritis (RA) is a chronic inflammatory autoimmune disease that is characterized by anarchic remodeling of joint architecture (11). Arthritogenic T cells and autoantibodies that migrate into the joints elicit inflammatory responses and lead to progressive destruction of cartilage and bone. Although the cause of RA is poorly understood, it is thought to be due to an adaptive immune response to autoantigen are accompanied by an inflammatory response.
Several groups have attempted to identify the role of Tregs in the pathogenesis of RA. A study using a collagen-induced arthritis model showed that depletion of Tregs increased susceptibility to the disease, whereas the other study indicated that Tregs were not involved in controlling proteoglycan-induced arthritis (12,13). The results in humans have been more diverse, since Tregs from patients with RA have been variously shown to be fully functional, partially defective, or fully defective (14-16). Therefore, the role of Tregs in RA is still a matter of debate.
K/BxN is a murine model of RA that has many of the clinical and histologic features of the human disease (17). In a previous study, we found that K/BxN mice contained normal number of Foxp3+ Tregs (18), and that these cells efficiently suppressed the proliferation of naïve CD4+ T cells, and cytokine production by effector CD4+ T cells, in vitro. Despite the existence of functionally intact Tregs, K/BxN mice develop autoimmune arthritis. This prompted us to investigate the role of Tregs in the pathogenesis of K/BxN arthritis. We generated Treg-deficient K/BxN mice (referred to as K/BxNsf mice hereafter) and examined diverse aspects of arthritis in these mice. We found that Treg deficiency in K/BxN mice resulted in earlier onset and more aggressive progression of arthritis, and that this coincided with increased activity of autoreactive T cells in secondary lymphoid organs and synovial tissue. Moreover, Treg deficiency increased the frequency and maturity of DCs in the inflamed synovia. Thus, our results suggest that although the activities of Tregs are not sufficient to block the development of arthritis in K/BxN mice, they reduce the severity of the autoimmune arthritis by lowering the activity of CD4+ T cells and DCs in secondary lymphoid organs and synovia.
MATERIALS AND METHODS
Mice
KRN TCR transgenic mice on a C57BL/6 background (K/B) were donated by Dr. Diane Mathis (17), and maintained in our facility. A cross between K/B and NOD mice generated transgenic progeny (K/BxN) and non-transgenic littermates (BxN). Female C57BL/6 mice bearing a scurfy (sf) allele (Jackson Laboratory, Bar Harbor, ME, USA) were backcrossed to NOD for more than 6 generations to generate NODsf congenic mice. A cross between female NODsf mice and male K/B mice generated K/BxN and K/BxNsf mice. All mice were maintained in a specific pathogen-free barrier facility at Hanyang University. The study was approved by institutional Animal Care and Use Committee.
Clinical and histopathologic evaluation of arthritis
The mice were inspected blind and disease severity was evaluated using a scoring system as previously described with modifications (19): 0, normal; 1, slight erythema and mild swelling confined to the mid-foot (tarsals) or ankle joint; 2, erythema and mild swelling extending from the ankle to the mid-foot; 3, erythema and moderate swelling extending from the ankle to the metatarsal joints; 4, intensive erythema and severe swelling encompassing the ankle, foot, and digits; 5, severe ankylosis of the ankle. In this system, the maximal score per mouse is 20 and scores are expressed as the mean arthritic index on a given day. The thickness of both hind ankles was measured axially across the malleoli using a caliper (Mitutoyo, Kanagawa, Japan). Hind paws were removed postmortem, fixed, and decalcified in Rapid-Cal Immuno solution (BBC, Stanwood, WA, USA). They were then embedded in paraffin, sectioned, and stained with H&E.
Flow cytometry
Single-cell suspensions of spleen, lymph node (LN) cells and synovial cells were stained with monoclonal Abs (mAbs) as previously described (18). The following mAbs were purchased from BD Biosciences (San Jose, CA, USA): anti-B220 (RA3-6B2), anti-CD4 (RM4-5), anti-IL-17 (TC11-18H10.1), anti-CD11c (R2-40), anti-CD11b (TC11-18H10.1), anti-CD40 (TC11-18H10.1), anti-CD86 (TC11-18H10.1), anti-CD3 (TC11-18H10.1), anti-NK1.1 (TC11-18H10.1), and anti-γδ TCR (B56). The following mAbs were obtained from eBioscience (San Diego, CA, USA): anti-CD80 (16-10A1), anti-CD25 (7D4), anti-Foxp3 (FJK-16s), anti-IFN-γ (XMG1.2), and anti-IL-4 (BVD6-24G2). The mAbs conjugated with FITC, PE, PerCP, or allophycocyanin were used.
Immunohistochemistry
Hind paw tissues were fixed, embedded in paraffin, and sectioned at 7 µm in thickness. Endogenous peroxidase was inactivated by treatment with 3% H2O2 in absolute methanol. After blocking nonspecific binding with normal goat serum in PBS, the sections were incubated with rabbit Ab to receptor activator of nuclear factor kappa-B (RANK; Santa Cruz Biotechnology, CA, USA) or RANK ligand (RANKL; Santa Cruz Biotechnology) at appropriate dilutions. The sections were incubated with secondary Ab and exposed to avidin-biotin-peroxidase complexes and 3,3'-diaminobenzidine (Dako, Carpinteria, CA, USA). The sections were counterstained with hematoxylin, and the samples were photographed with a photomicroscope (Olympus, NY, USA).
Synovial cell extraction
Synovial tissues from inflamed ankle joints of K/BxN and K/BxNsf mice were minced using scissor and digested with 0.5 mg/ml of Liberase Blendzyme II (Roche Diagnostics, Indianapolis, IN, USA) in plain RPMI 1640 medium for 45 min at 37℃. Digested tissue was washed three times in sterile plain RPMI and filtered through a cell strainer to collect single cell suspension.
Proliferation assay
Mouse glucose-6-phosphate isomerase peptide (GPI282-294; LSIALHVGFDHFE) was produced by Peprotech (Daejeon, Korea) (18). Splenocytes (105/well) from K/BxN or K/BxNsf mice were cultured in the presence of irradiated BxN splenocytes (105/well) as APC and the indicated concentrations of GPI282-294. After pulsing with 3H-thymidine (0.5 µCi/well; Amersham Biosciences, Piscataway, NJ, USA) for the last 9 hours of a total of 72 hours of culture, 3H-thymidine incorporation was assayed. All assays were conducted in triplicate.
RESULTS
Acceleration and exacerbation of arthritis in K/BxNsf mice
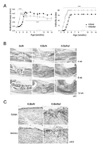
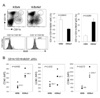
We found several differences between the arthritic symptoms of K/BxNsf mice, lacking Tregs, and their Treg-containing K/BxN littermates. The onset of arthritis occurred earlier and disease progression was accelerated in K/BxNsf mice (Fig. 1A). Ankle thickness, a measure of joint swelling, peaked 8 days earlier in K/BxNsf mice than in K/BxN mice. Despite the earlier decline of ankle thickness, joint deformity as indicated by arthritic indices and histological examination was much more severe in K/BxNsf mice than in the K/BxN mice up to 14 weeks of age (Fig. 1A and B). For example, the metatarsal bones were already invaded in the K/BxNsf mice at 4 weeks of age, while they were intact in K/BxN mice of the same age (Fig. 1B).
RANKL is a member of the TNF ligand superfamily, which is expressed by activated T cells and osteoblasts (20). RANKL binding to its unique receptor RANK activates the formation and function of osteoclasts, leading to bone resorption and loss. We performed immunohistochemical analyses to detect RANKL- and RANK-expressing cells in the hind paw joints of 6-week-old K/BxN and K/BxNsf mice. RANKL+ and RANK+ cells were much more abundant in the synovial tissues of the K/BxNsf mice than in those of the K/BxN mice (Fig. 1C). Thus, these results demonstrate that Tregs play a role in damping the severity of inflammation and bone destruction in the K/BxN arthritic model.
Treg deficiency enhances the quantity and activity of autoreactive CD4+ T cells
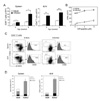
We examined whether Treg deficiency affected the number and/or activity of arthritogenic T cells. Total CD4+ T cell numbers were significantly higher in the spleens and joint-draining LNs (dLNs) of 4-8 week-old K/BxNsf mice than in their K/BxN littermates (Fig. 2A). These cells multiplied more than those from K/BxN mice even in the absence of exogenous GPI282-294 peptide, displaying enhanced self reactivity against endogenous GPI peptide (Fig. 2B).
We have shown previously that K/BxN mice contain numerous homeostatically proliferating CD4+ T cells retaining memory-like phenotypes, and that these cells are distinct from conventionally activated cells. Consistent with this, the large majority of CD4+ T cells populating the spleens and dLNs of K/BxN mice had memory (CD44hiCD62LloCD25-) phenotypes, while a few had undergone conventional activation (CD44hiCD62LloCD25+) or remained naïve (CD44loCD62Lhi) (Fig. 2C). The most prominent characteristic of the K/BxNsf mice was an increased frequency and absolute number of memory phenotype CD4+ cells in their spleens and dLNs (Fig. 2C and D). There were no comparable increase in conventionally activated CD4+ T cells in K/BxNsf mice (Fig. 2D).
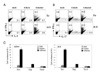
To determine whether Treg deficiency affects the development of effector helper T (Th) cells, we assessed the profiles of IFNγ-, IL-4-, or IL-17-producing Th1, Th2 and Th17 cells, respectively, in the spleen and dLNs of BxN, K/BxN and K/BxNsf mice. The K/BxNsf mice contained significantly more Th1 and Th2 cells in their spleens and dLNs than K/BxN mice (Fig. 3A and C), while the proportion of Th17 cells was lower than in K/BxN mice, even though there was no difference in the absolute number of Th17 cells. Immunofluorescence staining confirmed the existence of effector T cells in the spleens of K/BxN and K/BxNsf mice, especially in the extrafollicular areas but not in the follicular areas (Data not shown).
Thus our results show that Treg deficiency results in dramatic expansion of memory phenotype CD4+ T cells, presumably homeostatically proliferating T cells, in response to endogenous GPI self antigen. Treg deficiency also results in population of T cell zone by Th1 and Th2 cells but not Th17 cells. The latters seem not to be involved in the germinal center reaction.
Treg deficiency alters the cellularity of inflamed synovium
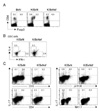
We examined whether Treg deficiency affects synovial cellularity in the K/BxN mice. To this end, we first tested whether Tregs are present in the synovial tissue of K/BxN mice. While Tregs were not seen in synovial tissue in the steady state, in the inflamed synovial tissue of K/BxN mice approximately 40% of the CD4+ T cells were Foxp3+ Tregs (Fig. 4A). Therefore, we suspected that Tregs play a substantial role in the pathology of the synovia.
To determine whether the absence of Tregs affects the homing of effector T cells to inflamed synovial, we examined the profiles of synovial T cells in K/BxNsf mice. As in the secondary lymphoid organs, K/BxNsf synovia contained more IFNγ+ Th1 cells and IL-4+ Th2 cells and fewer IL-17+ cells than K/BxN synovia (Fig. 4B). Since IL-17 can be produced by NK T cells and γδ T cells in addition to Th17 cells, we further characterized the phenotype of the IL-17-producing synovial cells. The majority of these cells in the arthritic joints expressed CD3 and γδ TCR, while few if any expressed CD4 or NK1.1 (Fig. 4C). Taken together, these results demonstrate that a deficiency of Tregs facilitates the influx of Th1 and Th2 cells into arthritic joints and does not affect the frequency of IL-17-expressing synovial γδ T cells.
Identification and characterization of DC subsets in synovial cells from K/BxNsf mice
Two major subsets of DCs, myeloid DCs (mDCs) and plasmacytoid DCs (pDCs), are abundant in the synovial fluid and perivascular regions of synovial tissues from patients with RA, suggesting that these DCs play an important role in inflammation. Hence, we tested whether Treg deficiency affected the DC subsets in the inflamed synovia. To this end, synovial cells extracted from arthritic joints of K/BxN and K/BxNsf mice were stained for CD11c, CD11b and B220. The proportion of CD11c+CD11b-B220+ pDCs was significantly higher and of CD11c+CD11b+B220- mDCs was slightly lower in K/BxNsf synovia than in K/BxN synovia (Fig. 5A). In addition, the expression levels of CD40 and CD86 were higher in the K/BxNsf pDCs than in those from K/BxN mice (Fig. 5B). The difference with regard to CD80 did not reach statistical significance. However, no upregulation of these molecules was seen in the mDCs of K/BxNsf mice (Data not shown). Thus, these results demonstrate that Treg deficiency leads to the accumulation of pDCs with higher costimulatory capacities in inflamed synovia.
DISCUSSION
Despite the existence of functionally intact Foxp3+ Tregs in K/BxN mice, these mice spontaneously develop arthritis. We were therefore prompted to consider the role of Tregs in these mice. We found that K/BxNsf mice lacking Tregs exhibited more serious symptoms of arthritis than their K/BxN littermates, including more extensive bone destruction, associated with increased proportion of RANKL+ cells in their synovial. They also contained more memory phenotype CD4+ T cells in their secondary lymphoid organs and synovia--- presumably proliferating homeostatically in response to autoantigens--- as well as more Th1 and Th2 cells. Importantly, pDCs with more mature phenotypes were more frequent in the synovia of the K/BxNsf mice than in those of the K/BxN mice. Thus, these results suggest that Tregs exert their effects on arthritogenic T cells and DCs, and so reduce the severity of disease.
We found that Treg deficiency resulted in general activation of T cells with the exception of Th17 cells. Unlike Th1 and Th2 cells, Th17 cells were more frequent in the K/BxN mice than the K/BxNsf mice. This result is intriguing in the light of the developmental and functional antagonism between Tregs and Th17 cells. We suspect that it may be due to the enrichment of Th1 and Th2 cells. Given that Th1 and Th2 cells can suppress the development of Th17 cells (21), such suppression may prevail over the direct effect of Treg deficiency. In addition, this result implies that Th17 cells do not play an important role in the exacerbation of arthritis in this model. Indeed, we observed that γδ T cells were more significant producers of IL-17 than Th17 cells.
We found above that Tregs, which are barely detected in synovia at steady state, became very numerous in inflamed synovia. This observation is in line with other reports showing enrichment of Tregs in inflamed tissues such as the synovial fluid of RA patients, pancreatic islets, colitis lesions, and the skin in graft-versus-host disease (22-25). This finding suggests that locally inflamed tissue provides a niche for Tregs to play a role in tissue-topical inflammatory responses. In this context, it would be of interest to further characterize the phenotypes and activities of synovial Tregs compared with those in secondary lymphoid organs.
An intriguing change in the Treg-deficient synovia was the increase in pDCs expressing elevated levels of CD40 and CD86. In agreement of this observation, others have detected the accumulation of pDCs in disease lesions in animal models and patients with autoimmune diseases, including type 1 diabetes, multiple sclerosis, Sjögren's syndrome and RA (26-29). Moreover, activated pDCs in local tissues preferentially secrete chemokines such as CXCL9, CXCL10, CCL3, CCL4 and CCL5, which can attract autoreactive T cells, so aggravating the disease (30-32). Similarly, we found that K/BxNsf synovia contained more Th1, Th2, and RANKL+ cells than K/BxN synovia. Therefore, we propose that synovial DCs are targets for the Treg activity that reduces synovial inflammation.
In conclusion, we have shown that Tregs reduce the severity of autoimmune arthritis by suppressing the activities of T cells and DCs in secondary lymphoid organs and synovial tissue.
 XML Download
XML Download