

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Although Th cells have the most effect on B cell maturation and differentiation once the B cells recognize protein antigens, it is increasingly clear that antigen presenting cells (APCs) directly affect B cells. APCs enhance B cell proliferation and differentiation (1-5). B cell-activating factor belonging to the TNF family (BAFF; also known as TALL-1, THANK, BLyS, and zTNF4), derived from dendritic cells (DCs) and macrophages, plays an important role in mediating such effects (6,7). Further, it is involved in B cell survival and maturation, germinal center formation, antibody production, and class-switching recombination (CSR) (8-10).
Activation-induced deaminase (AID) is indispensable for Ig CSR (11), and it has been demonstrated that BAFF induces AID expression (12). TGF-β1, a well-known switch factor for IgA and IgG2b isotypes (13-15), stimulates mouse macrophages and DCs to express BAFF (16), suggesting that TGF-β1 can regulate Ig synthesis indirectly.
Activin A, a member of the TGF-β1 superfamily, is the dimeric protein composed of b subunits linked by a disulfide bond, that include activin A (bAbA), activin AB (bAbB), and activin B (bBbB) (17). It has overlapping biological activities with TGF-β1 (18) and signals through Smad proteins (19). We have shown that activin A stimulates mouse B cells to express IgA isotypes (20). Nevertheless, it is not known if stimulatory activity of activin A on APCs is associated with B cell activation and differentiation.
In the present study, we explored the effects of activin A on BAFF expression by APCs such as DCs and macrophages to see if activin A regulates B cell differentiation indirectly by influencing APCs. We found that activin A can potentiate APCs to express BAFF through activation of the ALK4-Smad3 pathways.
MATERIALS AND METHODS
Reagents
Activin A, TGF-β1, GM-CSF and IL-4, anti-mouse BAFF antibodies were purchased from R&D Systems (Minneapolis, MN, USA). SB431542, SB203580, PD98059, and SP600125, ABTS were obtained from Sigma Chemical Co. (St. Louis, MO, USA).
Mice
BALB/c mice were purchased from Orient Co. Ltd (Gyeonggido, Korea) and maintained on an 8:16-h light:dark cycle in an animal environmental control chamber (Daehan Biolink. Co., Korea). Animals were fed Purina Laboratory Rodent Chow 5001 ad libitum. Eight- to twelve-week-old mice were used in this study. Animal care was in accordance with the institutional guidelines of Kangwon National University.
Culture of DCs and macrophages
The mouse dendritic cell line DC2.4 and the mouse macrophage cell line RAW264.7 were cultured in DMEM (2 mM Lglutamine; 100 U/ml penicillin; 100 g/ml streptomycin) plus 10% fetal bovine serum (HyClone Labs, Logan, UT, USA) in a humidified CO2 incubator. Bone marrow stem cells were isolated from BALB/c mouse femurs and cultured with GM-CSF (10 ng/ml) and IL-4 (10 ng/ml). After 7 days, appearance of DCs was determined with mouse anti-CD11c mAb (R&D System) by FACS. This procedure resulted in approximately 80% CD11c positive cell population. For the preparation of peritoneal macrophages, mice were injected with 3 ml of 4% thioglycolate broth in PBS intraperitoneally. After 72 h, peritoneal cells were harvested by peritoneal lavage with PBS containing 2% FBS, washed twice with HBSS, resuspended in 10% FBS-DMEM, and dispensed in 100-mm culture dishes. Cells were incubated in a CO2 incubator for 2 h at 37℃, and adherent cells were used as peritoneal macrophages.
Preparation of Plasmids
A mouse BAFF promoter (pBAFF) was cloned in our laboratory (16). The DN-Smad3 plasmid was provided by Dr. M. Kato (The Cancer Institute, Tokyo, Japan).
Transfection and luciferase assays
The reporter plasmids were co-transfected with the expression plasmids and pCMVβgal (Stratagene, La Jolla, CA) into DC2.4 cells by using a MicroPoratorTM MP-100 (Digital Bio Technology, Seoul, Korea); co-transfection was performed according to the manufacturer's instructions. Briefly, 2×106 cells were mixed with the reporter plasmid and the expression plasmid and pulsed twice at 1000 V for 30 ms. Luciferase and β-galactosidase assays were performed as described (21).
RT-PCR
RNA preparation, reverse transcription, and PCR were performed as described (21). PCR primers were synthesized by Bioneer Corp. (Seoul, Korea). The primers for mouse BAFF were forward primer 5'-GCC GCC ATT CTC AAC ATG AT-3' and reverse primer 5'-TTA GGG CAC CAA AGA AGG TG-3' (product size: 468, 409 bp). All reagents for RT-PCR were purchased from Promega (Madison, WI, USA). PCR was also performed with β-actin to allow for normalization of cDNA concentrations in each set of samples. PCR products were resolved by electrophoresis on 2% agarose gels.
ELISA for mouse BAFF
ELISA was performed as described previously (16). Anti-mouse BAFF antibody (2µg/ml) was added to 96 well microplates. After incubation overnight at 4℃, the plates were washed and blocked with 1% gelatin for 1 h. Supernatant samples (50µl) or standard protein (mouse recombinant BAFF) diluted in 0.5% gelatin were added to the wells. After incubation for 1 h at 37℃, the plates were washed again, and 2µg/ml monoclonal anti-mouse BAFF antibody was added for 1 h at 37℃. The plates were then washed and incubated with goat anti-rat IgG-HRP for 1 h. After washing, 0.2 mM ABTS was added to the wells, and 10 min later, the colorimetric reaction was measured at 405 nm with a VERSAmax ELISA reader (Molecular Devices, Sunnyvale, CA, USA).
RESULTS AND DISCUSSION
Effect of activin A on BAFF expression by mouse DCs
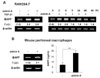
APCs are involved in B cell activation and differentiation (4,5) and BAFF is one of the important factors expressed by APCs such as macrophages and DCs (1,22,23). Since we have demonstrated that TGF-β1 stimulates mouse macrophages to express BAFF (16), it was worthwhile to investigate if activin A, a member of the TGF-β superfamily, can modulate APCs to express BAFF. To explore the possibility that activin A regulates BAFF expression in mouse DCs, we first examined the effect of activin A on the levels of endogenous BAFF transcripts. Activin A induced BAFF transcription in the mouse dendritic cell line, DC2.4. Optimal induction was observed under conditions of 10 ng/ml activin A and 9-h stimulation (Fig. 1A). Similarly, activin A increased the promoter activity of BAFF (Fig. 1B). Since activin-A increased the BAFF transcription and promoter activity, it was necessary to determine its effect at the protein level. Two types of BAFF are known to exist in a secreting and a membrane-bound form (22,7). Here, activin A increased the secretion of BAFF by DC2.4 for at least 48 hrs following stimulation (Fig. 1C). Concentration of activin A (i.e., 10 ng/ml), which significantly increased BAFF expression, little affected cell growth as shown in Fig. 1D. These results indicate that activin A actually regulates mouse DCs BAFF gene expression. Further, activin A also stimulated BAFF expression at the mRNA and protein levels in primary bone marrow-derived DCs (Fig. 2). Taken together, our results clearly show that activin A stimulates mouse DCs to express BAFF.
Effect of activin A on BAFF expression by mouse macrophages
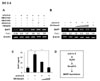
We proceeded to determine if activin A regulates BAFF expression in mouse macrophages. As shown in Fig. 3A, activin A significantly increased BAFF transcription in RAW264.7, a mouse macrophage cell line. BAFF transcript was detectable as early as 3 h after stimulation (Fig. 3A). These results are largely consistent with our early finding that TGF-β1 induces BAFF in such cell line (16). In primary peritoneal macrophages, activin A also stimulated BAFF expression at both the transcriptional and protein levels (Fig. 3B). These results reveal that, in mouse, activin A not only stimulates DCs but also macrophages to express BAFF.
Involvement of the Smad pathway in activin A-induced BAFF expression
We subsequently investigated the mechanisms underlying activin A-induced BAFF expression. Since the activin A signal activates not only ALK4/Smad pathway but also mitogen-activated protein kinases (MAPKs) pathway, such as p38MAPK, JNK, and ERK (19,24-26,27), we asked the involvement of ALK4/Smad pathway and MAPK pathway in activin A-induced BAFF expression. SB431542 (an ALK4 inhibitor) completely abrogated activin A-induced BAFF transcription, whereas PD98059 (ERK inhibitor), SP600125 (JNK inhibitor), and SB203580 (p38 inhibitor) had little effect (Fig. 4A), suggesting that activin A induces BAFF expression through the ALK4 but not MAPKs pathway. Further, activin A-induced BAFF transcription was abrogated by DN-Smad3 in a dose-dependent manner (Fig. 4B). This was paralleled by changes in BAFF expression at the protein level (Fig. 4C). Taken together, these results indicate that ALK4-Smad3 pathway is mainly responsible for activin A induced BAFF expression (Fig. 4D).
In summary, we demonstrate in the present study that activin A stimulates mouse APCs to produce BAFF and that this is attributed to ALK4-Smad3 signaling. BAFF is believed to be the most important macrophage- and DC-derived B cell-activating factor (1). Among functions of BAFF, it can activate B cells to express AID gene (28,29). We have recently demonstrated that macrophage-derived BAFF induces AID expression in mouse B cells (30). Therefore, these in vitro studies imply that activin A causes APCs to produce BAFF, which may have important effects on the Ig isotype switching in vivo.


 XML Download
XML Download