

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Human cytomegalovirus (HCMV) is a ubiquitous herpesvirus that infects 50~85% of adults. Primary infection with HCMV always leads to the virus persisting for long periods in the host as a latent infection. In immune-compromised individuals, HCMV can reactivate and may give rise to a clinical illness (1). Several studies show that bone marrow progenitors act as a reservoir of HCMV and transmit the viral genome to peripheral blood monocytes, without lytic-gene expression, until they leave the circulation and undergo tissue-specific differentiation to macrophages (2-6). Infection and disease by HCMV are major complications of allogeneic bone marrow transplantations (BMT) (7). HCMV infection commonly occurs one to three months after transplant and may be followed by disease manifestations approximately 15 days later (8).
Dendritic cells (DCs) are potent antigen presenting cells that bridge innate and adaptive immunity (9). Positioned at interfaces between organism and environment and in lymphoid tissues, DC are uniquely suited to recognize and acquire pathogens, process macromolecular antigens into peptides, and initiate immune responses in naive T cells, including CD4+ Th cells and CD8+ cytotoxic lymphocytes (CTL). There is considerable interest in harnessing the potent antigen presenting power of DC for patient benefit, and DC-based vaccine trials are ongoing.
Expression of HCMV antigens in DCs can be achieved by transfection with cDNAs or infection with viruses encoding HCMV antigens (10). However, this technique may have some constrains, including gene delivery, prolonged gene expression, efficiency, and toxicity (11). We have explored transduction of protein antigen into DCs as an alternative to genetic modification to elicit CTL in vitro. Bacterial recombinant proteins containing the 11-amino acids HIV TAT protein transduction domain (PTD; YGRKKRRQRRR) translocate across cell membranes in vivo and in vitro (12,13). The pp65 (65 kDa lower matrix phosphoprotein) (14), known as glycoprotein 64 or UL83 of a virion tegument protein and the main component of the enveloped subviral particle is an immunodominant target of CD4+ as well as CD8+ T cell responses to HCMV (15).
Our results demonstrated that DCs pulsed with N- and C-terminal pp65 protein fused with Tat (Tat/pp65 N&C) enhanced induction of pp65-specific CTL in vitro.
MATERIALS AND METHODS
Cloning
The pp65 cDNA (UL83; HCMV AD169 accession number NC001347) was obtained from viral DNA. Primers corresponding to the 5' and 3' ends of pp65 cDNA were cgaactcgagATGGAGTCGCGCGGTCGCCGT, with an XhoI restriction site, and gcgcgcggatcc TCAACCTCGGTGCTT, with a BamH I restriction site, respectively. The purified HCMV-pp65 gene was digested with XhoI and BamH I, and then subcloned into a pET15b expression vector (Novagen, Darmstadt, Germany) which had been digested with the same restriction enzymes.
Next, Tat/pp65 gene was annealed to generate a promoter encoding 11 amino acids (HIV-1 Tat47-57: YGRKKRRQRRR) from the basic domain of HIV-1 Tat. The sequences are, at the 5' and 3' ends, respectively: gcagcatatg-TATGGAAGGAAGAAGCGGAGACAGCGACGAAGA-ctcgag-ATGGAGTCGCGCGGTCG and gcgcgcggatcc-TCAACCTCGGTGCTT. The NdeI-BamH I-digested Tat/pp65 gene was ligated into the NdeI-BamH I-digested pET15b vector.
A minigene was constructed encoding the N-terminal pp65, pp65N and C-terminal pp65, pp65C using PCR extension with the following primers, at the 5' and 3' ends, respectively: pp65N forward and reverse primers -cgaactcgag-ATGGAGTCGCGCGGTCGCCGT, gctggatcc-TCAATTTTTGGGACACAACAC and pp65C forward and reverse primers -gaactcgag-ATGATAATCAAACCGGGCAAG; gcgcgcggatcc-TCAACCTCGGTGCTT.
The XhoI-BamH I-digested pp65N and pp65C gene was cloned into the XhoI-BamH I-digested pET15b or pET15b-Tat vectors and finally the DNA sequence can be confirmed by automated DNA sequencing (model 373A; Applied Biosystems, Inc., CA, USA).
Production and purification of recombinant protein in E.coli
The BL21 transformed with pp65/pET15b plasmid was inoculated into 2×500 ml of LB broth containing 100µg/ml ampicillin. After the cells had grown until OD600=0.5~0.7, IPTG (Isopropyl-β-D-thiogalactoside, Duchefa Co., Netherland) was added to a concentration of 1.5 mM, and the incubation was continued for an additional 3 h. The cells were harvested, and a 40 ml binding buffer (8 M Urea, 100 mM NaCl, 20 mM HEPES, pH 8) was added and sonicated (model VC130; Sonics & Materials, Inc., CT, USA).
The recombinant pp65 was purified under denaturing conditions. To denature pp65, harvested cells were disrupted by sonication in a binding buffer containing 8 M urea. After centrifugation, supernatants containing pp65 were immediately loaded onto a 5 ml Ni2+-nitrilotriacetic acid Superflow (QIAGEN, Germany) column. The column was washed with 2 volumes of binding buffer, and 8 volumes of a washing buffer (8 M Urea, 100 mM NaCl, 20 mM HEPES, 10 mM imidazole, pH 8). For refolding step, the column was washed with urea in each 1 volume of refolding buffer (0.2% Tween20, 0.5% Glycerol, 0.5 M NaCl in PBS, pH 7.4) at decreasing concentration (6, 4, 2, 1, 0.5, 0.2, and 0.1 M). The fusion protein was eluted with an elution buffer (1 M imidazole, 0.1% Tween20, 1% Glycerol, 0.5 M NaCl in PBS, pH 7.4). Dialysis was then performed to remove residual salt.
To confirm the presence of purified protein, we performed 12.5% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. The protein was visualized either by Coomassie blue staining. The protein concentration in fractions was estimated by the Bradford procedure using bovine serum albumin as a standard (16).
Induction of monocyte-derived dendritic cells and LCL
Six healthy laboratory volunteers were included in this study. Consent forms and approval for this study were obtained from the donors and the Institutional Review Board of The Catholic University of Korea, College of Medicine. HLA typing was performed in Hematopoietic Stem Cell Bank (Catholic medical center, Seoul, Korea).
PBMC were obtained from peripheral blood by standard Ficoll-Paque Plus (Amersham Biosciences, Piscataway, NJ, USA) separation. The adherent cells were isolated in RPMI 1640 complete medium containing 5% FBS at 107 cells/well in 6-well plates following 2 h incubation at 37℃. The non-adherent cells were removed and the adherent cells were cultured for 7 days in RPMI 1,640 complete medium containing 100 ng/ml GM-CSF (Endogen, Woburn, MA, USA) and 500 U/ml IL-4 (Genzyme, Cambridge, MA, USA). Fresh culture medium was added on day 3. After 7 days, non-adherent and loosely adherent cells were collected by vigorous pipetting. More than 90% of the cells showed DC phenotype (MHC class I molecule+, class II molecule+, CD1a+, HLA-DR+, CD14-, CD80+) by FACS (data not shown).
Immature DCs (1~2×106 cells/ml) were treated with recombinant protein for 2 h at 37℃ and then recultured for 1 day in the presence of 200 units/ml tumor necrosis factor-α (TNF-α, R&D System, Inc., Minneapolis, MN, USA), 1µg/ml prostaglandin (PG)-E2 (Sigma-Aldrich Corp, St. Louis, MO, USA) and 1µg/ml lipopolysaccharide (LPS, Sigma-Aldrich) for maturation.
LCLs were generated with concentrated supernatants of the B95-8 cell line [16]. LCLs were also transduced with pLXSN-pp65 retrovirus (LCL/pp65), to permit selection of a pure population for target cells (19). Both LCLs and LCL/pp65 were maintained in RPMI 1,640 (GibcoBRL, Grand Island, NY, USA) supplemented with 10% fetal bovine serum (FBS, GibcoBRL), 2 mM L-glutamine (GibcoBRL) and antibiotic antimycotic mixture 1X; JBI, Daegu, Korea).
Isolation of CD4+ and CD8+ T cells
CD4+ and CD8+ T cells were separated from PBMC using MACS microbeads and MidiMACS positive selection column type LS according to the manufacturer's instructions (Miltenyi Biotec, Bergisch-Gladbach, Germany). 108 PBMC were co-incubated with 50µl of anti-CD4 or anti-CD8 microbeads for 30 min at 4℃. Then cells were washed with 2 mM EDTA in PBS and applied onto the LS column placed in a magnetic filed. Negative cells were passed through the column by rinsing with 2 mM EDTA in PBS. Then the column was removed from the magnetic filed and positive cells were flushed out. The purity of the sorted CD4+ or CD8+ T cells was measured by FACS and was >93%.
IFN-γ ELISPOT assay
The effector cells (PBMCs, 106 cells/well) and DCs pulsed with each protein (105 cells/well) were mixed and incubated 4 hr at 37℃ in 96-well U-bottomed plate (Falcon, Becton Dickinson Labware, Franklin Lakes, NJ, USA). Control wells contained effector cells alone and effector cells in the presence of mock DCs. After that, a 1 : 50 dilution of effector cells (104 cells/well at final volume of 200µl) were added to a 96-well anti-IFN-γ-coated ELISPOT plate (AutoImmun Diagnostika [AID], Strassberg, Germany) in duplicates after blocking with RPMI 1,640 complete medium (10% FBS, 15~20 min, 37℃).
The cells were incubated for 18~20 h at 37℃ in 5% CO2 and washed six times with washing buffer according to the manufacturer's instructions. A secondary biotinylated anti-IFN-γ antibody was then added at 100µl/well for 2.5 hr. Plate was washed five times with buffer, and was incubated with streptavidin-horseradish peroxidase for 2 h at room temperature. Plates were washed with washing buffer and spot color was developed by adding 3-amino-9-ethylcarbazol substrate diluted in acetate buffer containing H2O2. Plates were then washed with distilled water to stop the reaction. After drying, images of the wells were acquired using the AID ELISPOT software version 2.1 (AID). All the experiments were run at least in duplicate depending on the number of cells available. The results are shown as the number of spot-forming cells (SFC)/106 cells.
In vitro CTLs generation
PBMCs (106 cells/ml) were cocultivated with autologous DCs pulsed with each protein in 24-well plate (Falcon) in RPMI 1,640 complete medium containing 10% FBS. DCs were exposed to 25 Gy of γ irradiation before use as stimulator cells. The CTL culture was stimulated weekly following a regimen of decreasing responder: stimulator ratios from 20 : 1 at day 0, 10 : 1 at day 7 over period of 2 weeks.
Cytotoxicity assay
Cytotoxic activity of the generated CTL was assessed 1 week after the second stimulation by 51Cr release using autologous LCL, LCL/pp65, and allogeneic (HLA-nonmatched) LCL/pp65 as target. All target cells were labeled with 100µCi 51Cr in 0.2 ml RPMI 1,640 complete medium containing 10% FBS at 37℃ for 1 h. After three washes, the target cells were counted and seeded in triplicate in 96-well V-bottomed plates (Falcon) at 5×103 cells per well. The effector cells were then added at an indicated effector-to-target (E/T) ratio. After centrifugation at 1,200 rpm for 2 min, the plates were incubated at 37℃ for 4 hr. The supernatant fluids were harvested and 51Cr content was measured. The percentage of specific incorporation was calculated as: [(cpm test-cpm spontaneous)÷(cpm maximum-cpm spontaneous)]×100.
Tetramer staining
The Phycoerythrin (PE)-labeled HLA-A2402 tetramers complexed with the peptide, QYDPVAALF (amino acids 328-336 of the HCMV pp65 protein) was obtained from Proimmune (Oxford, UK). Cells were incubated with a PE-labeled HLA-A2402/pp65 328-336 tetramer and fluorescein isothiocyanate (FITC)-labeled anti-CD8 antibody (BD Biosciences) for 1 h at 4℃. After incubation, the cells were washed and fixed in PBS with 1% paraformaldehyde. Samples were analyzed by FACS and at least 50,000 events were collected for each sample.
RESULTS
Characterization of recombinant TAT/pp65 fusion protein
Six different recombinant pp65 proteins were produced for this study: pp65, pp65 fused with Tat, N-terminal pp65 (pp65N), C-terminal pp65 (pp65C), N-terminal pp65 with Tat (Tat/pp65N) and C-terminal pp65 with Tat (Tat/pp65C). The structure of each protein was schematically shown in Fig.1A. The amount of each protein produced from 1 liter of E.coli. was 1.25 mg of pp65, 0.35 mg of Tat/pp65, 1.2 mg of pp65 N, 2 mg of Tat/pp65N, 16 mg of pp65C, and 7.2 mg of Tat/pp65C (Fig. 1B). The purified proteins were shown to have a molecular mass of ~34 kDa for the pp65 N and pp65C, ~35 kDa for the Tat/pp65 N and Tat/pp65C, ~65 kDa for the pp65, and ~66 kDa for the Tat/pp65 by Coomassie blue staining (Fig. 1C). We used mixture of Tat-pp65N and Tat-pp65C or pp65N and pp65C protein to generate pp65-specific CTL because yield of production of full pp65 protein was low.
Transduction efficacy of Tat/pp65 fusion protien into DC
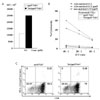
To analyze that Tat fusion protein can effectively transfer into DCs, DCs were incubated with 0.01, 0.1 and 1 uM of Tat/GFP or GFP for 2 h, following maturation for 1 day and GFP expression in DCs was examined by FACS. When DCs was incubated with Tat/GFP, high expression of GFP protein was observed (Fig. 2A).
We investigated whether transduction of recombination fusion protein affects death of DCs using Annexin V/7-AAD staining. Incubation of DCs with pp65 N&C and Tat/pp65 N&C protein did not trigger cell death including apoptosis (Fig. 2B).
Enhancement of T cell stimulation by DC pulsed with Tat/pp65 fusion protein
To measure the responses of T cells to the recombinant pp65 protein, PBMCs were stimulated with DCs pulsed with various concentration of pp65N&C or Tat/pp65N&C, and the frequency of IFN-γ producing cells was measured by ELISPOT assay. Protein concentration of 0.01 and 0.1µM of Tat/pp65 N&C showed higher frequency of IFN-γ producing cells compared with that of pp65N&C (Fig. 3A). Because protein concentration of 0.1µM of Tat-pp65 N&C showed the highest frequency of IFN-γ producing cells, 0.1µM of protein was used in this study.
To discriminate CD4+ and CD8+ T cells responses, CD4+ and CD8+ T cells were incubated with DCs pulsed with pp65 N&C and Tat/pp65 N&C protein and then, frequency of IFN-γ producing cells was examined by ELISPOT assay. CD8+ T cells as well as CD4+ T cells showed higher frequencies of IFN-γ producing cells when was stimulated with DCs pulsed with Tat/pp65 N&C protein compared with pp65 N&C (Fig. 3B). The pp65-specific CD8+ T cells recognized Peptide NV9 (HLA-A2 restricted) reported as CTL peptides. Frequency of IFN-γ producing cells increased 1.6 fold in CD8 T cells, and 0.5 fold in CD4 T cells in Tat/pp65N&C protein group, indicating that DCs pulsed with Tat/pp65 N&C were more potent in stimulating CD8+ T cells than DCs pulsed with pp65 N&C.
In vitro generation of pp65-specific CTLs by Tat/pp65 fusion protein
After PBMCs were stimulated with DCs pulsed with pp65 N&C or Tat/pp65 N&C protein three times, we produced pp65-specific CTL in vitro and investigated that it could target LCL pulsed with pp65 N&C or Tat/pp65 N&C protein using IFN-γ ELISPOT assay. T cells generated by DCs pulsed with Tat/pp65N&C protein showed higher frequencies IFN-γ producing cells compared with that of by DCs pulsed with pp65N&C protein, when stimulated with LCL pulsed with pp65 N&C protein (Fig. 4A).
The cytotoxic activity of the generated CTL was assessed by 51Cr release assay using autologous (HLA-A24, 33) and allogeneic (HLA-A02, 26) LCL, LCL/pp65N&C as targets (Fig. 4B). Cytotoxic activity was significantly higher against autologous LCL pulsed with pp65N&C in T cells generated with DCs pulsed with Tat/pp65 N&C compared with T cells generated by DCs pulsed with pp65 N&C (60% versus 43% at E/T ratio 40 : 1). There was no significant cytotoxic activity against autologous LCL unpulsed with pp65N&C, allogenic (HLA-nonmatched) LCL unpulsed and pulsed with pp65N&C as targets.
For tetramer assay, tetrameric Q9F-HLA-A2402 complex was used and the percentage of A24/pp65 tetramer+ CD8+ T-cells was analyzed by FACS. A24/pp65 tetramer+ CD8+ T-cells showed 0.48% in Tat/pp65 N&C and it was twice higher percentage than 0.23% in pp65 N&C (Fig. 4C). Together, these results suggest that DCs pulsed with Tat/pp65 could effectively induce pp65-specfic CTLs.
DISCUSSION
The vaccine for treatment of HCMV aims at the activation and expansion of antigen-specific CTL through the presentation of MHC class I -restricted eptiopes to CD8+ T cells. DCs have been the potent antigen presenting cell to priming CTL and interest has been focused on the way to deliver tumor antigen to DCs. Although several epitopes of pp65 for pulsing to DCs have been reported until now (20), pp65 epitopes that can be recognized by T cells for HCMV immunotherapy remain to be limited. Peptide generates antigen-specific immune response, but is the short of half time of peptide/MHC complexes and MHC restriction that limits this approach to patients with specific MHC allotypes. The DCs pulsed with pp65 protein can induce CTL against various epitopes because protein antigen contains the undefined antigens including multiple class I-restricted epitopes as well as helper peptide. In addition, antigen protein can be produced on a large scale with low cost. Development of system to deliver protein antigen to DCs including Tat fusion protein will improve availability of protein antigen.
To demonstrate whether fusion of pp65 to Tat would enhance the immunogenic properties for T cells stimulation, pp65N&C and Tat/pp65N&C protein was produced and compared with immune responses each other. DCs pulsed with pp65N&C or Tat/pp65 N&C protein directly stimulated PBMC and CD4+ and CD8+ T cells (Fig. 3B). In addition, pp65-specific CTL could be generated in vitro with DCs pulsed with pp65N&C or Tat/pp65N&C protein (Fig. 4). DCs pulsed with Tat pp65N&C protein markedly enhanced the generation of pp65-specific CTL in vitro. One of mechanism to improve immune responses of Tat fusion protein may be the increased uptake and expression of Tat fusion protein by DCs as shown in Fig. 2A, because cation of Tat protein and anion on plasma membrane have the binding ability by ionic interaction (21, 22).
Although DCs pulsed with Tat/pp65N&C showed the higher frequencies of IFN-γ producing cells compared with DCs pulsed with pp65N&C in 0.01 and 0.1 uM concentration, it exhibited the decreased frequency in 1 uM of protein concentration (Fig. 3A). High protein concentration of Tat protein may be affect on the viability of DCs. Tat/pp65N&C showed the higher increase rate in CD8+ T cells responses than CD4+ T cells responses (Fig. 3B), indicating that Tat fusion protein is involved with cross presentation of DCs.
In conclusion, fusion of Tat protein to pp65 protein in this study enhanced immunogenicity of pp65 antigen and DCs pulsed with Tat-pp65 fusion protein induced effective pp65-specific CTL. These results will be helpful for the development of adoptive T cell immunotherapy and DC-based immunotherapeutic anti-viral vaccines for the treatment of a broad variety of immune-suppressed patients after bone marrow transplantation.



 XML Download
XML Download