

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The leukocyte common antigen (CD45), the most common hematopoietic lineage marker, belongs to a family of transmembrane-type protein tyrosine phosphatases with high molecular masses of 180 to 220 kDa (1-7). Using many monoclonal antibodies (mAbs) against CD45, it was revealed that CD45 comprises 5~10% of lymphocyte surface proteins, known as one of the most abundant glycoproteins expressed on lymphocytes (1).
Charbonneau et al. observed a significant sequence similarity between the tandem repeats in the cytoplasmic domains of two proteins, CD45 and protein tyrosine phosphatase (PTP) 1B (8). Subsequent cloning of CD45 at the cDNA and genomic levels revealed several interesting characteristics about the primary structure of this molecule (9-11). The extracellular domain of human CD45 varys in length (391~552 amino acids) depending on which combination of exons are alternatively used to form the CD45 mRNA. The three alternatively used exons of CD45, exons 4, 5, and 6, encode peptide segments designated A, B, and C, respectively. In human, five different isoforms of CD45 mRNAs have been isolated, which contain all three exons (ABC isoform), two of the three exons (AB and BC isoform), only one exon (B isoform), or no exons (O isoform), respectively (9-11). All of the isoforms (schematically shown in Fig. 1) have the same 8 amino acids at their amino-terminus, which are followed by the various combinations of A, B, and C peptides (66, 47, and 48 amino acids long, respectively). The remaining regions (the 383-amino-acid extracellular region, the 22-aminoacid transmembrane peptide, and the 707 amino-acid-cytoplasmic region) have the identical sequences in the all isoforms. The N-terminal region of CD45 is known to be heavily glycosylated (12). Therefore, alternative mRNA splicing of CD45 can result in a significant degree of heterogeneity in the extracellular domain due to differential O-linked glycosylation as well as the structure changes of the molecule.
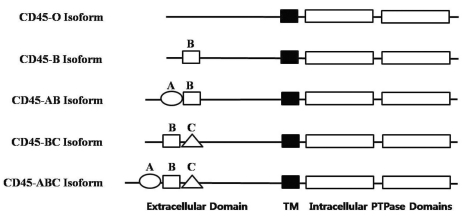
As a result of the variability of the N-terminal region of CD45, mAbs raised against the CD45 protein recognize either all of the CD45 isoforms (CD45 mAb), or only a subset of the isoforms ("restricted" CD45R mAb). Thus, the suffix RA, RB, or RO indicates the requirement of the amino acid residues corresponding to exon A (RA), exon B (RB), or a lack of amino acid residues corresponding to exon A, B and C (RO) for the CD45 epitope expression, respectively. Accordingly, CD45 mAb binds to all isoforms, whereas CD45RA mAb binds to ABC and AB isoforms, CD45RB mAb binds to ABC, AB, BC, and B isoforms, and CD45RO mAb binds only to the 180 kDa isoform, which lacks any of the alternatively used exons (O isoform). In this report, we analyzed the characteristics of seven murine mAbs raised against the human leukocyte common antigen (CD45) (AP4, DN11, SHL-1, YG27, P1, P6, and P14). By using these antibodies, we were able to not only immunoprecipitate the CD45 isoforms, but also differentiate cellular expression profiles of the isoforms. In addition, by transiently transfecting COS-7 cells with the plasmids expressing CD45 isoforms, we examined the binding specificities of the mAbs to those isoforms.
Go to : 
MATERIALS AND METHODS
Production of monoclonal antibody
Seven murine mAb (AP4, DN11, SHL-1, YG27, P1, P6, and P14) were developed by immunizing Balb/c mice with various native human cellular immunogens. Briefly, AP4 was raised by immunizing PHA-activated human peripheral blood mononuclear cells (PBMC), and other antibodies were raised with cells from various origins: thymic stromal cells (for DN11), Jurkat (for SHL-1), thymocytes (for YG27), and resting human PBMCs (for P1, P6, P14). After 6 week-old Balb/c mice were immunized i.p. with each immunogen (107 cells), the spleens were removed, and 108 spleen cells were fused with 107 SP2/0-Ag14 mouse myeloma cells using polyethylene glycol (PEG 4000, Rahway, NL). The hybrids were cultured in flat-bottom microculture trays at 37℃ in an atmosphere of humidified air conditioning 5% CO2, and selected in HAT media. After 10 days, culture supernatant was harvested and tested for reactivity to human lymphocytes by indirect immunoflurescence method using flow cytometry. Seven of the resulting hybridoma clones, whose supernatant was reactive to human lymphocytes, were named AP4, DN11, SHL-1, YG27, P1, P6, and P14. Cells from a microculture well were subcloned by limiting dilution, and the culture supernatant of the clones was tested for antibody production. For production of the mAb in ascites form, pristane-treated Balb/c mice were injected i.p. with 5×106 hybridoma cells, and the ascites were collected after 1 week of injection. mAb was purified from ascites.
Determination of antibody isotype
The isotype of AP4, DN11, SHL-1, YG27, P1, P6, and P14 were determined by enzyme immunoassay using mouse monoclonal subtyping kit EK-5050 (Hyclone, Utah, USA). Isotyping was performed with rabbit anti-murine isotype-specific antisera (IgG1, IgG2a, IgG2b, IgG3, IgM, Kappa, Lambda) followed by peroxidase-labeled goat anti-rabbit IgG as the secondary antibody. with the addition of o-phenylene diamine and hydrogen peroxide substrate, positive samples turned an intense yellow.
Cells and cell lines
We screened the expression of the AP4, DN11, SHL-1, YG27, P1, P6, and P14 antigen on the freshly isolated peripheral blood erythrocytes, lymphocytes, monocytes, granulocytes and platelet from the healthy volunteers. The tumor cell lines were purchased from American type culture collection (atcc, Rockville, MD) and cultured in RPMI-1640 medium supplemented with 10~20% fetal calf serum.
Flow cytometry
Fresh cell suspension from patients and healthy donors were examined for the AP4, DN11, SHL-1, YG27, P1, P6, and P14 antigen expression by flow cytometric analysis using AP4, DN11, SHL-1, YG27, P1, P6, and P14 mAb. Cells (106) were incubated with saturating amounts of the purified AP4, DN11, SHL-1, YG27, P1, P6 and P14 or a control immunoglobulin for 30 min at 4℃. Thereafter, cells were washed two times with phosphate-buffered saline (PBS), incubated with fluorescein isothiocyanate (FITC)-conjugated goat anti-mouse as secondary antibody for 20 min at 4℃ and washed two times with PBS. Cells were analyzed on a FACS (COULTER, USA). Samples containing over 20% AP4, DN11, SHL-1, YG27, P1, P6, and P14-positive cell were regarded as positivity.
Immunohistochemical study
We screened tonsil and thymus obtained from the surgical pathology files of Chungbuk National University Hospital. All tissues had been fixed in 10% formalin and embedded in paraffin, and indirect immunoperoxidase technique and antigen retrieval method by microwave heating were employed (13,14).
Chromogen used as 3,3'-diaminobenzidine. Counterstain was not performed and the reaction pattern was analyzed based on serial hematoxylin-eosin stained sections. We defined positive cells by staining pattern along cell membrane.
Immunoprecipitation
Fresh cell suspensions of tonsil from patient of chronic tonsillitis and of thymus from patient of congenital heart disease were lysed in lysis buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1% NP-40 and 1 mM PMSF [phenyl methyl sulfonyl fluoride]). Lysates were precleared successively with protein A-Sepharose and rabbit anti-mouse IgG bound to protein A-Sepharose at 4℃ for 4 h. 20µl of seven mAb coupled to protein A-Sepharose were used for immunoprecipitation with precleared cell lysates at 4℃ overnight. Immune complexes were washed with a buffer containing 0.25% NP-40, 5 mM PMSF, 10 mM Tris, pH 8.0, 150 mM NaCl, 5 mM KI, and 5 mM EDTA. After extensive washing, the immunoprecipitates were eluted by boiling for 5 min in SDS sample buffer and analyzed on 6% SDS-PAGE under non-reducing condition, with appropriate molecular weight markers. After electrophoretic transfer of immune complex, the nitrocellulose was blocked with 5% skimmed milk in Tris-buffered saline (10 mM Tris-HCl, 150 mM NaCl, pH 7.6) containing 0.05% Tween-20 (TBST). The bound peroxidase was visualized using the ECL chemoluminescence detection system (Amersham).
Production of tansfectants
The cDNAs encoding the five CD45 isoforms, which were inserted into the cloning site of a slightly modified version of the pcDL-SRα296 expression plasmid (15), termed pSP65-SRα2. The resultant plasmids were used to transiently transfect the COS-7 cells by the DEAE-dextran method (16). COS-7 cells were harvested by trypsination and were seeded into 10 cm plate at a final density of 1×106 cells/plate. After 12 h, the cells were washed twice with prewarmed phosphate-buffered saline (PBS) (138 mM NaCl, 2.6 mM KCl, 10 mM Na2HPO4, 1.8 mM KH2PO4, pH 7.4) and were fed with transfection mixture (DME, 0,25 mg/ml DEAE dextran, 0.1 M Tris, pH 7.3 containing DNA), using 4 ml/plate for 12 h. After 12 h, the cells were washed with prewarmed PBS and were incubated with 10% DMSO reagent at room temperature. 3 min after addition of the 10% DMSO reagent, the medium was removed by aspiration and the monolayers were washed with prewarmed PBS. The cells were subsequently incubated in culture medium containing 0.1 mM chloroquin for 2 h 30 min. The cells were washed with prewarmed PBS, and incubated with fresh culture medium in CO2 atmosphere at 37℃ for 48 h. Resulting cells express each of the CD45 isoforms. The binding specificities of seven mAb were characterized by flow cytometric analysis using the COS-7 cells that expressing the individual human CD45 isoforms. The anti-CD45/FITC (T29/ 33; DAKO, Denmark) was used as a positive control. Samples containing over 10% AP4, DN11, SHL-1, YG27, P1, P6, and P14-positive cell were regarded as positivity.
Go to :
RESULTS
Determination of antibody isotype
The isotype of AP4, DN11, SHL-1, YG27, P1, P6, and P14 mAb were determined by enzyme immunoassay. The isotype of AP4 was IgG2b whereas the isotypes of the other remaining mAbs were all categorized into IgG1.
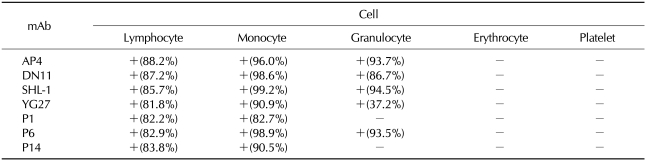
Reactivity of AP4, DN11, SHL-1, YG27, P1, P6, and P14 to hematopoietic cells
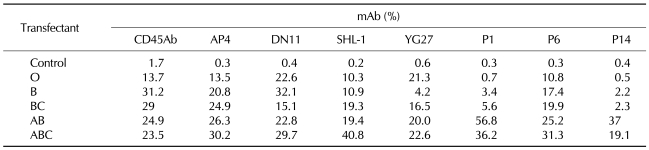
To evaluate the recognition profile of the mAbs in various hematopoietic cells, normal human hematopoietic cells from peripheral blood were analyzed by flow cytometry. Five monoclonal antibodies (AP4, DN11, SHL-1, YG27, and P6) were reactive with lymphocytes, monocytes, and granulocytes, but not with erythrocytes or platelet. Two monoclonal antibodies (P1 and P14) were reactive with lymphocytes and monocytes, but not with granulocytes, erythrocytes, or platelets (Table I).
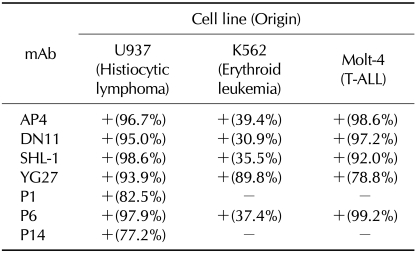
Reactivity of AP4, DN11, SHL-1, YG27, P1, P6, and P14 to various leukemic cell lines
Similarly to the above experiment, we examined the recognition profile of the mAbs in leukemic cells, U937 (Histiocytic lymphoma), K562 (Erythroid leukemia) and Molt-4 (T-ALL) cells. AP4, DN11, SHL-1, YG27, and P6 mAb were reactive with all of the tested leukemic cell lines. P1 and P14 were reactive with U937, but not with K562 or Molt-4 (Table II).
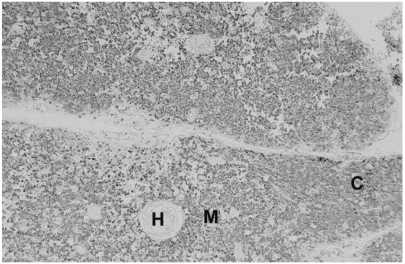
Reactivity to formalin fixed thymus and tonsil
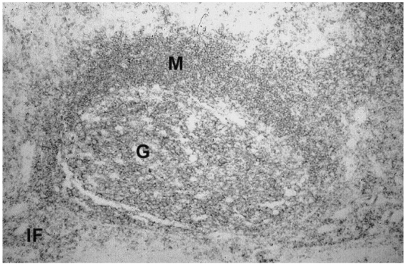
To validate their clinical usage, the immunoreactivity of mAbs on formalin-fixed paraffin-embedded human tissue was checked. All the seven mAbs displayed strong membranous immunoreactivity in nearly all cases of immune cells in formalin fixed paraffin embedded thymus (Fig. 2) and tonsil (Fig. 3) samples.
Characterization of seven mAb target antigen
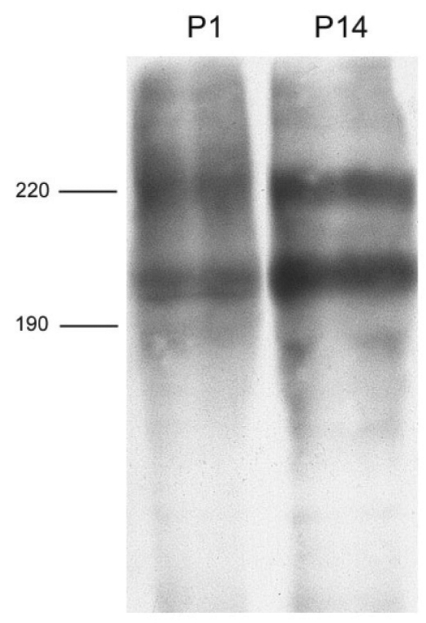
To identify the antigen immuniprecipitated with these seven mAbs, lysates of thymus and tonsil were analyzed. The molecular weights of proteins recognized by AP4, DN11, SHL-1, YG27, and P6 were examined by immunoprecipitation using thymus lysates. AP4 recognized two proteins of 220 and 205 kDa whereas DN11, SHL-1, YG27, and P6 recognized three proteins of 220, 205, and 190 kDa (Fig. 4). The molecular weight of the antigen recognized by P1 and P14 was determined by immunoprecipitation using tonsil lysates. Two discrete bands of the antigen corresponded to two molecules of approximately 225 and 200 kDa (Fig. 5).
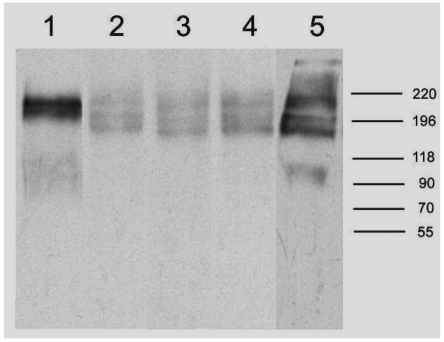 | Figure 4Immunoprecipitation of thymus lysates with AP4, DN11, SHL-1, YG27 and P6 mAb. Lane 1; AP4 lane 2; DN11 lane 3; SHL-1 lane 4; YG27 lane 5; P6. Cells were lysed in lysis buffer containing 1% NP-40. precleared cell lysates were immunoprecipitated with AP4, DN11, SHL-1, YG27 and P6 mAb. The immunoprecipitates were analyzed by SDS-PAGE (6%) under nonreducing condition.
|
Transfection study
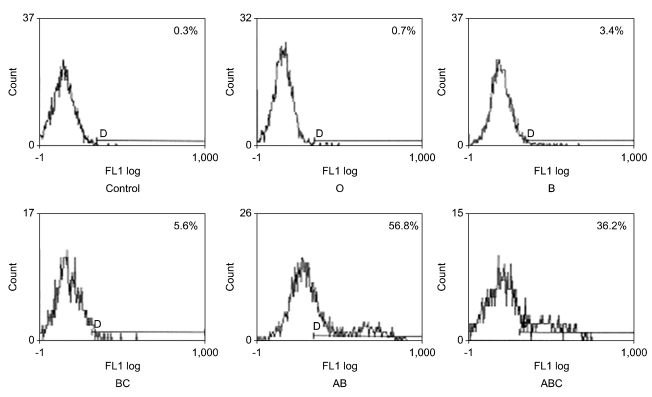
To confirm which isoform of CD45 is recognized by each of these seven mAbs, transfecton study was performed. In transfection study, four mAb (AP4, DN11 (Fig. 6), SHL-1, and P6) bound to all five isoforms (the ABC isoform, AB isoform, BC isoform, B isoform and O isoform), one mAb (YG27) bound to four isoforms (the ABC isoform, AB isoform, BC isoform and O isoform) and two mAb (P1 (Fig. 7) and P14) bound to two isoforms that include exon A encoded sequences (the ABC isoform and AB isoform). In these results, we confirmed that AP4, DN11, SHL-1, YG27 and P6 are conventional CD45 mAb and P1 and P14 are CD45RA mAb (Table III).
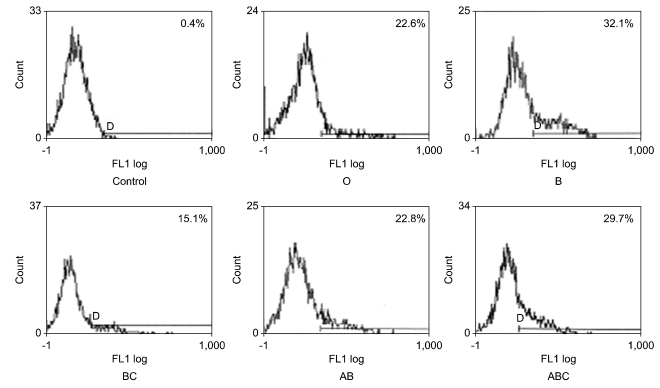 | Figure 6Transfection study of DN11 mAb. Flow cytometric analysis of DN11 using COS-7 cells expressing the individual human CD45 isoforms. DN11 bound to all five isoforms (the ABC isoform, AB isoform, BC isoform, B isoform and O isoform).
|
Go to :
DISCUSSION
In this report, we examined the specificities of seven mAbs raised in our lab by using transiently transfected COS-7 cells that express each of five CD 45 isoforms. Based on the binding patterns, the seven mAbs are classified into two groups: AP4, DN11, SHL-1, YG27, and P6 are CD45 mAbs whereas P1 and P14 are CD45RA mAbs. Because of their abundant expression of hematopoietic cells, we were easily able to obtain mAbs against CD45 by immunizing conventional cellular immunogen, such as thymocytes, PBMCs and leukemic cell lines, but mAb against CD45RB or CD45RO could not be obtained.
Based on the expression profiles of CD45 mAbs that were previously studied and reported, we classified the generated mAbs by their flow cytometric patterns. In flow cytometric analysis of the seven mAb with hematopoietic cells, five monoclonal antibodies (AP4, DN11, SHL-1, YG27 and P6) were reactive with lymphocytes, monocytes and granulocytes but not with erythrocytes and platelets (Table I). Two monoclonal antibodies (P1 and P14) were reactive with lymphocytes and monocytes but not with granulocytes, erythrocytes and platelets (Table I). According to Leukocyte Typing V (17), CD45RB mAb is reactive with erythrocytes and CD45RA mAb is not reactive with granulocytes. Therefore, in terms of CD45 antigen expression pattern recognized by seven mAbs, there is no CD45RB mAb at least and we supposed that P1 and P14 should be a CD45RA mAb. U937 (Histiocytic lymphoma), K562 (Erythroid leukemia). Molt-4 (T-ALL) were analyzed by flow cytometry. AP4, DN11, SHL-1, YG27 and P6 mAb were reactive with these all leukemic cell lines. P1 and P14 only were reactive with U937, but not with K562 and Molt-4 (Table II). According to Leukocyte Typing V (17), CD45RO mAb is not reactive with U937. Therefore, there is no CD45RO mAb among seven mAbs. CD45RA mAb was not reactive with K562 and Molt-4. Therefore, those data also support that P1 and P14 would be a CD45RA mAb. In terms of molecular weight analysis AP4, P1, and P14 were immunoprecipitated with two proteins of 220 and 205 kDa (Fig. 5). But DN11, SHL-1, YG27 and P6 recognized three proteins of 220, 205, and 190 kDa (Fig. 4). Above expression profiles and molecular results also support that AP4, DN11, SHL-1, YG27 and P6 would be a conventional CD45mAb and P1 and P14 would be a CD45RA mAb.
In order to confirm those findings, we transiently transfected COS-7 cells with each of 5 isoform cDNA constructs of CD45, and examined the specificities of the seven mAbs against the isoforms. Although the percentage of the positive ones against these mAbs among the transfected COS-7 cells appeared to be relatively low, we were able to classify their specificities by comparison of commercially available anti-CD45 Abs with known specificity (Table III). Of the seven mAbs tested, four mAb (AP4, DN11 (Fig. 6), SHL-1, and P6) recognized a sequence common to all of the five isoforms (ABC, AB, BC, B, and O) (Table III). One mAb (YG27) recognized four isoforms (ABC, AB, BC and O), and two mAb (P1 (Fig. 7) and P14) recognized isoforms that include exon A encoded sequences (ABC and AB) (Table III). Thus, these results indicate that AP4, DN11, SHL-1, YG27 and P6 are anti-CD45 mAbs whereas P1 and P14 are anti-CD45RA mAbs. YG27 mAb unusually revealed negative reactivity to COS-7 cells transfected with B construct, but remaining four constructs revealed positive reactivity we categorized YG27 mAb as a CD45 mAb.
Although the development of various CD45 mAbs has been previously reported, only a few antibodies against CD45RB and CD45RO were successful. As expected, the majority of the mAbs that we developed recognized all isoforms, and only two mAbs (P1 and P14) recognized both ABC and AB isoforms. None of mAbs recognizes B and O isoforms. It is because the epitopes required for the generation of CD45RO or CD45RB mAbs might be heavily glycosylated, which process is essential for the biological function of thymocytes, lymphocytes and leukemic cells (17-20). In case of using CD45 antigens overproduced in E. coli, antibodies recognizing only the epitope composed of the polypeptide backbone will be produced, resulting in failure of antibody production that recognize the natural structure of the antigen. Accordingly, we utilized whole immune cell extract for immunization to produce practical antibodies for diagnostic or therapeutic usage in this study, but the generation of CD45RO or CD45RB mAbs was not successful.
In immunohistochemical study, all the seven mAb showed strong membranous staining patterns in case of all the hematopietic cells in formalin-fixed paraffin-embedded thymus and tonsil samples (Figs. 2 and 3). This result indicates that these mAbs may be used singly or together for the detection and the differential diagnosis of hematopoietic malignancies in surgical pathology (13,14).
Several key issues related to the physiological role of CD45 isoforms remain mostly unsolved, such as the identification of its extracellular ligands as well as intracellular substrates, and the mechanism responsible for regulation of CD45 catalytic activity, albeit a few limited published data (17-21). These questions are currently being investigated using a wide range of technical approaches and cell systems, which seems to be more advanced by the development of various CD45 mAbs. Currently, the modulation of CD45 function by mAbs has been suggested as an optional treatment to control autoimmune diseases, transplant rejection, and even cancer (22-25). Therefore, the possibility of therapeutic significance of these seven mAbs should be evaluated in the near future.
Go to :
 XML Download
XML Download