

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
T cells can be classified into three separate classes: naïve, effector and memory. Antigen-specific naïve CD8 T cells exist at very low frequencies. But during an immune response, these cells undergo massive clonal expansion and finally form memory T cells. Memory T cells are well suited to combat pathogens because they are present at higher numbers than naïve CD8 T cells, persist for extended periods due to antigen- independent homeostatic turnover, and can mediate a substantially accelerated recall response to secondary infection (1,2). Thus, many efforts have been made to elucidate how to efficiently elicit greater numbers and longer-lived memory T cells.
It has been proposed that a brief period of antigen encounter can commit naïve cells to a program of effector and memory differentiation (2-6). However, it has been shown that the duration and strength of antigenic and costimulatory signals can also regulate the fate of developing effector and memory T cells (7,8). In this case, memory cells are proposed to differentiate from naïve cells by virtue of qualitative and quantitative differences in the perceived signal. In particular, early inflammatory cytokine signals known as "third signal" have been shown to be involved in the induction of effector response and the generation and maintenance of effector and memory cells (9-14). However, the exact mechanisms regulating the fate of activated T cells still remain undefined. Previously, we showed that IL-12 priming during antigenic stimulation dramatically increased the population of effector and memory CD8 T cells mainly by preventing activation-induced cell death (9), suggesting that development of memory cells could be regulated by early IL-12 signaling.
CD43 is a cell-surface sialoglycoprotein expressed by a variety of hematopoietically derived cells, including T lymphocytes (15-17). Posttranslational modifications result in two glycoforms of CD43: 115 kDa and 130 kDa. The mAb 1B11 recognizes the activation-associated glycoform (130 kDa) of CD43 (18,19). The 130 kDa glycoform of CD43 bears two core O-glycans, an oligosaccharide structure that can be created by the action of the core β-1-6-N-acetylglucosaminyltransferase. 1B11 binding is low on naïve CD8+ T cells and high on Ag-specific effector CD8+ T cells, and becomes reduced again on Ag-specific memory CD8+ T cells (20). Although CD43 is one of the most abundant T cell surface glycoproteins, its function is still debated. It has been suggested to have anti-adhesive functions in T cell trafficking and homing (21-23). Some evidence suggests that CD43 provides a highly charged barrier to cell-cell interaction (24,25). A topological view of T cell-APC interactions has been proposed (26), suggesting that CD43 may be a cell surface regulatory protein that damps T cell responses by its physical presence (27). This hypothesis could be supported by the fact that CD43 is excluded from the immunological synapse during T cell activation (27-29). Additionally, CD43 has been also shown to play a role in T cell activation. Some studies showed that CD43 transduces multiple activating signals that ultimately induce expression of certain genes during T cell activation (30-32) and plays a costimulatory role in vitro and in vivo (33-35). In contrast, other groups have shown that CD43-/-T cells are hyperproliferative in vitro (21,36).
In the present study, we analyzed the mechanisms by which IL-12 priming contributes to the activation and the enhanced survival of CD8+ T cells and observed dramatically decreased expression of CD43 in activated CD8+ T cells primed by IL-12. To determine the role of CD43 expression in the survival of activated CD8 T cells, we purified CD43lo and CD43hi cells after IL-12 priming and analyzed the function and survival of each population. CD43lo effector CD8+ T cells exhibited reduced cytolytic activity, lower granzyme B expression and reduced IFN-γ production but showed significantly increased survival both in vivo and in vitro compared to CD43hi cells. These CD43lo effector CD8 T cells are associated with higher expression of CD62L than CD43hi effector CD8 T cells. Together, these results suggest that the expression of the activated form of CD43 is significantly down-regulated by IL-12 priming, which gives rise to a preferential, long-lived CD8+ T cell memory population that is partly associated with the levels of CD62L expression.
MATERIALS AND METHODS
Mice
Female C57BL/6 mice were purchased from The Charles River Japan (Shizuoka, Japan). OT-I TCR transgenic mice were purchased from The Jackson Laboratory (Bar Harbor, ME). All mice were housed under specific pathogen-free conditions and were used between 6 and 12 weeks of age following institutional animal care and use committee protocols.
Antibodies and reagents
All antibodies were purchased from BD Bioscience-Pharmingen (San Diego, CA), unless specified otherwise. Recombinant human IL-2 and murine IL-12 were purchased from R&D Systems (Minneapolis, MN). CD8α+ T cell isolation kits and anti-PE microbeads were obtained from Miltenyi Biotec (Auburn, CA).
In vitro T cell activation, isolation, adoptive transfer and infection
Spleen cells from OT-I TCR transgenic mice were stimulated with OVA257-264 peptide (SIINFEKL; referred to as OVAp) in complete IMDM supplemented with 2 mM L-glutamine, 50µM 2-ME, and 10 U/ml human rIL-2 in the presence of rIL-12 (5 ng/ml). After in vitro stimulation, the CD8+ T cells were purified by negative selection using magnetic bead separation (MACS) according to the manufacturer's instructions (Miltenyi Biotec). CD43lo and CD43hi cells were then purified by negative selection and positive selection using anti-CD43-PE and anti-PE microbeads. Purified CD43hi and CD43lo CD8+ T cells (2×106 in 200µl of PBS) were transferred into naïve C57BL/6 mice via tail vein injection. For recall response, 1×105 purified CD43hi and CD43lo CD8+ T cells in 200µl of PBS were transferred into naïve C57BL/6 mice via tail vein injection, and at day 1 after transfer, the mice were intranasally challenged with 2×107 pfu of recombinant adenovirus expressing OVA (rAd/OVA).
Surface staining, intracellular staining, and flow cytometric analysis
In order to count the total number of donor T cells, recipient mice were sacrificed, and cells from spleens were resuspended in FACS buffer (1% FBS, 0.03% sodium azide in PBS) at a concentration of 1×107 cells/ml. A total of 100µl of these cells (1×106 cells) was stained for CD8 (clone 53-6.7), CD43 (1B11), CD62L (MEL-14) or CD127 (A7R34), and samples were acquired on FACS Calibur™ (BD Biosciences, San Jose, CA). PE or APC-conjugated OVA-specific MHC I tetramer, Kb/OVA-Tet, was produced as described elsewhere (9), and the optimal concentration was determined by titration. Cells were stained for 40 min at 4℃ using fluorochrome-conjugated Abs and Kb/OVA-Tet, washed, and fixed in PBS containing 2% formaldehyde before analysis by flow cytometry. For intracellular staining, purified CD43hi and CD43lo cells were co-cultured at 37℃ for 5 h with OVA peptide and BFA. After culture, the cells were first stained for surface markers, then washed, fixed and permeabilized with FACS buffer containing 0.5% saponin (Sigma-Aldrich, Seoul, Korea). The cells were then stained with anti-IFN-γ (XMG1.2). For granzyme B staining, cells were fixed and permeabilized with FACS buffer containing 0.5% saponin after surface marker staining. Cells were stained with anti-granzyme B (GB12) or isotype control antibody conjugated to PE (Caltag Laboratories). Gates were set on lymphocytes by forward and side scatter profiles, and the data were analyzed using CellQuest™ Pro (BD Biosciences), FlowJo™ software (Windows version 5.7.2, TreeStar, San Carlos, CA), and WinMDI version 2.8 software (The Scripps Research Institute, La Jolla, CA).
Detection of apoptotic death
Apoptosis of in vitro activated T cells was determined by Annexin V and 7-amino-actinomycin D (7-AAD) staining, as recommended by the manufacturer (BD Bioscience, San Diego, CA). In brief, purified CD43hi and CD43lo CD8 T cells were seeded in 24-well flat-bottom plates (1×106 cells/well) with IL-2 in 2 ml of complete IMDM. At the indicated time points, these cells were washed, resuspended in Annexin V binding buffer at a concentration of 1×106 cells/ml, incubated with Annexin V-FITC and 7-AAD, and analyzed by flow cytometry.
Cell-mediated cytotoxicity assay
MACS-separated CD43lo and CD43hi effector cells were used in the CTL assay in vitro. Peptide-pulsed EL4 target cells (1×104/well) were added to serial dilutions of effector cells (prepared as described above) in 96-well round-bottom plates at E:T cell ratio of 1:1 to 20:1. After 4 h at 37℃, cytotoxicity was quantified by measurement of cytosolic lactate dehydrogenase (LDH) in the culture supernatant (n=3) using cytotoxicity detection kit (Roche Diagnostics). Specific lysis for each E:T cell ratio was expressed as: specific lysis=[(experimental release)-(spontaneous release)/(target maximum-target spontaneous release)]. Spontaneous LDH release in the absence of CTL was <10% of the maximal cellular release by detergent lysis. All experimental procedures and assays were performed two or more times with similar results.
RESULTS
CD43 expression regulated by initial IL-12 priming during primary CD8+ T cell stimulation
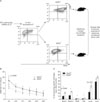
Previously we showed that IL-12 priming during initial antigenic stimulation significantly enhanced the survival of activated CD8 T cells and increased the memory cell population after adoptive transfer (9). Recent studies have suggested that activation markers, such as CD43 and CD27, define distinct subpopulations of memory CD8+ T cells that differ in their capacities to mount recall responses (20,37). To determine whether IL-12 priming induces any changes in CD43 expression during in vitro stimulation, we analyzed CD43 expression on OT-I cells stimulated with cognate OVA (SIINFEKL) in the presence or absence of IL-12. The level of CD43 and CD44 on naïve cells was low, but after antigenic stimulation, most OT-I cells expressed CD44hi activated phenotype and some of them started to express CD43 on day 1 (Fig. 1). There was no significant difference in the CD43 expression between non-primed and IL-12-primed cells during this period. However, beginning on day 3, IL-12-primed cells showed a relatively lower proportion of CD43-positive cells than non-primed cells, and this selective down-regulation of CD43 by a subpopulation of effector cells was observed clearly on days 3, 5 and 7 (Fig. 1A). To determine whether CD43 is associated with the typical characteristics of memory precursors, we assessed CD43 expression together with other activation markers such as CD127 (38), KLRG-1 (39) and CD62L (40,41) that identify memory precursors. There was no significant difference in the CD127 expression between CD43hi cells and CD43lo cells (Fig. 1B). However, CD43lo cells showed a relatively higher expression of CD62L than CD43hi cells (Fig. 1B). These data suggest that IL-12 priming during antigenic stimulation induced lower CD43 expression than in non-primed cells and that this CD43lo phenotype among activated T cells may be associated with enhanced survival of activated CD8 T cells.
Enhanced survival of CD43lo cells induced by IL-12
CD43 is one of the most abundant T cell surface glycoproteins. But the function of CD43 as a regulatory protein in T-cell activation remains unclear. To define the role of CD43 down-regulation by IL-12 priming, we analyzed the survival potential of IL-12-primed CD43hi and CD43lo CD8 T cells by sorting the two populations by MACS and then adoptively transferring equal numbers into B6 recipient mice (Fig. 2A). We analyzed the "take" of donor cells in recipient mice at various time points after adoptive transfer. At all time points, ~1.5-2-fold higher levels of CD43lo donor cells were detectable in the peripheral blood (Fig. 2B). Some studies suggest that CD43 plays a role in the trafficking of T cells to sites of inflammation (23,42). In addition, a study using CD43-deficient mouse showed that CD43 negatively regulates T lymphocyte homing to secondary lymphoid organ (22). Thus, it was possible that the higher level of CD43lo donor cells in the blood was caused by differential homing to lymphoid organs. We therefore examined the trafficking of CD43hi and CD43lo donor CD8 T cells to the lymphoid organs and non-lymphoid organs in the recipient mice. At day 7 after adoptive transfer, CD43lo donor cells gave rise to 2-fold higher percentages of engraftment in all lymphoid organs examined such as spleen and mesenteric and inguinal lymph nodes and 1.5-fold higher percentages in non-lymphoid organs such as lung and liver (Fig. 2C). These data exclude the possibility that increased frequency of CD43lo donor cells in the peripheral blood was due to differential trafficking to various organs.
Next, we investigated whether the phenotypic distinction of activated CD8 T cells by CD43 affected the susceptibility to apoptosis. To this end, activated OT-I cells were sorted to CD43hi and CD43lo cells and each subset was rested with IL-2 and monitored for apoptotic cell death at various time points (Fig. 3). CD43lo T cells showed a similar death rate to CD43hi T cells by 48 h into the resting period, but exhibited a significant reduction in apoptotic death by 72 h (p<0.001). The proliferation of CD43lo effector cells was similar to that of CD43hi effector cells (data not shown). These results were consistent with the report that CD43-/- mice showed decreased death of Ag-specific CD8 T cells (35). Taken together, our results indicate that the phenotypic change in CD43 expression induced by IL-12 was associated with reduced cell death and the subsequent enhanced survival of effector and memory CD8 T cells.
Functional difference between CD43lo and CD43hi cells induced by initial IL-12 priming
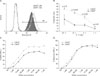
As previously stated, the reported positive and negative roles of CD43 in T cells are inevitably opposed to each other. Thus, we investigated whether the CD43lo and CD43hi T cell subsets exhibited any difference in effector functions. To this end, the subsets of CD43hi and CD43lo cells were sorted after antigenic stimulation in the presence of IL-12, and granzyme B expression and killing activity were measured (Fig. 4). A comparison of these effector properties between CD43lo and CD43hi T cells demonstrated that CD43lo CD8 T cells contained less granzyme B than CD43hi CD8 T cells (Fig. 4A), and exhibited significantly less killing of OVA peptide-pulsed targets (Fig. 4B). These results were consistent with the report that CD43 up-regulation shows a strong correlation with the acquisition of effector function (20). It is possible that CD43 expression affects functional avidity of CD8 T cells since CD43 can provide a charge barrier based on the topological view of cell-cell interactions (24). It is reported that TCR avidity of CD8 T cells to peptide/MHC complex correlates with ex vivo cytotoxicity and IFN-γ secretion (43). So, we investigated whether CD43lo and CD43hi phenotype affected the functional avidity of OVA-specific effector CD8 T cells by measuring dose-responsive IFN-γ production. CD43hi effector cells exhibited higher IFN-γ expression than CD43lo effector cells in all peptide concentrations used (Fig. 4C). But when the data are normalized to % maximum response, dose-responsive IFN-γ production of CD43hi and CD43lo effector cells was comparable (Fig. 4D). This indicates that functional avidity of CD43hi effector cells and CD43lo effector cells was similar. Together, these results suggest that the CD43lo phenotype induced by IL-12 priming was associated with lower granzyme B expression, killing activity and IFN-γ secretion but not with TCR avidity.
One of the functional memory properties is recall response to the same antigen. So, we next examined recall responses of adoptively transferred CD43lo and CD43hi populations by challenging Ag. To test recall proliferation of memory cells generated from CD43hi and CD43lo donor cells, we adoptively transferred equal numbers of sorted CD43hi and CD43lo CD8 T cells into naïve mice and challenged them with recombinant adenovirus expressing OVA (rAd/OVA) 1 day after adoptive transfer. We picked this early time point of challenge since CD43hi and CD43lo subsets displayed differential survival rates even a few days after transfer. We challenged mice with the rAd/OVA virus intranasally because the distribution of CD43lo and CD43hi donor T cells in the lung tissues was similar. Most of the CD8 T cells detected in the lungs at the peak (day 5 after challenge) were donor OT-I cells, and the number of OVA/Tet+ cells among lymphocyte-gated mononuclear cells were 3~5 fold higher in mice that received CD43lo cells than in mice that received the equal number of CD43hi cells (Fig. 5). These results are consistent with the report that CD43lo CD8 T cells mediate stronger recall responses than CD43hi cells (37).
Enhanced survival of CD43lo CD8 T cells partly associated with CD62L expression
As shown in Fig. 1B, CD43lo cells showed a relatively higher expression of CD62L than CD43hi cells. High CD62L expression among CD8 T cell memory population has been shown to be a phenotypic characteristic of the central memory subset (44). To determine whether the higher expression of CD62L was involved in the enhanced survival of CD43lo cells, we analyzed the memory potential of IL-12-primed CD62Lhi and CD62Llo CD8 T cells by MACS purification and a subsequent adoptive transfer experiment. After sorting, the ratio of CD43hi:CD43lo cells among CD62Lhi population was approximately 1:9 while that among CD62Llo population was ~3:7 (Fig. 6A). At various time points after adoptive transfer, CD62Lhi donor cells gave rise to 1.2-fold higher levels of surviving memory cells in peripheral blood (Fig. 6B). At day 32 after transfer, CD62Lhi donor cells increased to 2-fold higher levels in lymph nodes, compared with CD62Llo donor cells, while there was no significant difference between CD62Lhi cells and CD62Llo cells in other tissues including spleen, lung and liver (Fig. 6C). These data demonstrate that the enhanced survival of IL-12-primed CD43lo CD8 T cells is partly associated with CD62L expression.
DISCUSSION
Despite progress over the last few years in elucidating the mechanisms of memory cell development in T cell immunity, it is still not well understood. The presence of proinflammatory cytokines in addition to TCR and costimulatory signals can have a profound effect on the outcome of immune responses. Our previous study suggests that initial IL-12 signaling strongly influences the programming of memory T cell development (9). In this study, we have identified a new surface marker, CD43, the expression of which is regulated by initial IL-12 signaling. CD43 expression was previously shown to be a marker that distinguishes between memory and effector T cells (18,20,35). Expression of CD43 is relatively low on naive CD8 T cells but high on Ag-specific effector CD8 T cells, and becomes reduced again on Ag-specific memory CD8 T cells (20). Our data indicate that expression of the activated form of CD43 (1B11) is down-regulated by IL-12 priming (Fig. 1A), and all adoptively transferred CD43lo and CD43hi cells eventually changed into CD43lo cells in the recipient mice. But these antigen-specific CD8 T cells exhibited CD43hi phenotype again when they met the same antigen (data not shown). What are the functional consequences of the changes in CD43 expression on activated T cells? Recent studies suggest that activation markers such as CD27 and CD43 define three distinct subpopulations of memory CD8 T cells that differ in their capacities to mount a recall response (37). By adoptively transferring purified CD43hi and CD43lo T cells, we have shown that CD43lo cells induced by IL-12 priming have enhanced survival (Fig. 2B) and mediate stronger recall responses (Fig. 5). Although all adoptively transferred CD43lo and CD43hi cells eventually turned into CD43lo cells in recipient mice, the CD43lo subset showed higher survival and recall potential than CD43hi subset. In addition, we found a strong correlation between decreased expression of CD43 on antigen-specific CD8+ T cells and decreased functions such as granzyme B, cytolytic activity and IFN-γ secretion. Until now, the exact role of CD43 has not been well defined due to its seemingly contradictory roles in several processes such as cell adhesion and costimulation (20,22,35,45). Our results support the idea that CD43 is promiscuously related to T cell activation, effector functions, survival and recall responses but not T cell homing. The previous works have shown that TCR signaling leads to the redistribution of CD43 outside of the mature immunological synapse, and that relocalization of CD43 is required for optimal T cell activation (29,46). In addition, CD43 appears to provide intracellular signals that synergize with those of the TCR (47). These previous reports and our results both support the idea that CD43hi cells have both TCR and CD43 signals at the same time, consequently CD43hi cells, which have a stronger signal, may exhibit more effector function than CD43lo cells. The progressive T cell activation model proposes that signal strength regulates the progression of the T cell through the hierarchy of proliferation, differentiation and death (7). Thus, it is likely that down-regulation of CD43 by IL-12 priming diminishes the overall strength of T cell stimulation, which provides a distinct advantage for the survival of activated CD8 T cells.
In summary, the data presented here suggest that the effect of IL-12 priming on the prolonged survival of activated CD8 T cell is mediated by down-regulation of CD43 expression. And decreased expression of CD43 has the potential to regulate generation and survival of effector/memory CD8+ T cell. Although further studies are needed to characterize the exact mechanisms of CD43 in the generation of memory CD8+ T cells, our findings suggest that a decreased expression of CD43 on activated CD8+ T cells by IL-12 priming is involved with enhanced generation of central memory precursors.




 XML Download
XML Download