

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The interest of researchers in medicinal plants as natural sources of active principles has markedly increased over the past decades with particular attention paid to the polysaccharide compounds of various traditional Asian medicines (1,2). For example, Cordyceps militaris (CM) has been used as a folk medicine in Korea, China, and Japan for a long time. Its chemical components, including cordycepin (3,4), nucleoside, and various polysaccharides (5,6), have been widely studied. However, little is known about its immune-enhancing effects or the effects of its mechanism on murine APCs.
Antigen presenting cells, especially Dendritic cells (DCs), play a critical role in the initiation of immune responses and the induction of immune tolerance. DCs are the most important accessory cells for the activation of naïve T cells and the generation of primary T cell responses (7). They can acquire and process antigens in the periphery, and migrate to secondary lymphoid tissues where they prime primary T cell responses. Since T cells can only recognize antigens presented on MHC molecules, modulation of MHC-restricted antigen processing pathways may provide novel pharmacological targets for the regulation of T cell responses.
In the present study, we examined the effects of Cordyceps militaris water extract, CME, on the in vitro function of DCs with the goal of characterizing the mechanism of CME as an immune-enhancing agent. We used OVA as an exogenous antigen in conjunction with CME, and then compared the change in cross presentation of CME-related DCs to that of a control group along with the level of MHC class I and II molecules. Further, we examined the immune-enhancing activity of CME on peritoneal macrophages. We found that CME activated several parameters of macrophage activation such as nitric oxide and cytokine expression and antigen presenting to T cells. Ultimately, CME may explain some of the therapeutic efficacies of CM, which has been used in folk medicine to treat various diseases including cancer.
MATERIALS AND METHODS
Reagents
Lipopolysaccharide (LPS) was purchased from Sigma (St. Louis, USA). Cell culture media DMEM, antibiotic-penicillin/streptomycin solution, and fetal bovine serum (FBS) (Hyclone, Logan, USA) were used for the cell culture. Cordyceps militaris water extract (CME) were supplied by CM Biotec. (Kangnung, Korea) (6).
Cell culture
T cell hybridomas, CD8OVA1.3 and DOBW (8), were kindly provided by Dr. Clifford V. Harding (Case Western Reverve University, Cleveland, OH). The DC cell line (DC2.4) was obtained from the Dana-Farber Cancer Institute, Boston, MA, USA (9). CD8OVA1.3, DOBW, and DC2.4 cells were maintained in DMEM medium supplemented with 10% FBS, 100 U/ml of penicillin, 100µg/ml of streptomycin, and 2 mM L-glutamine. Bone marrow-derived DCs were generated from BALB/C mice bone marrow precursor cells (10). Bone marrow cells obtained from femurs of mice were cultured in a 6-well plates in a culture medium supplemented with 200 U/ml rmGM-CSF. At days 3 and 4 after initiation of the culture, nonadherent cells were discarded by replacing the culture medium with fresh medium containing cytokines, after gentle shaking. DCs were harvested by gentle pipetting at day 6. Primary macrophages were collected from the peritoneal cavities of mice (8-week-old male ICR mice) after an intraperitoneal (i.p.) injection of 3 ml of 3% thioglycolate broth (Sigma, St. Louis, MO) 4 days before harvesting. The peritoneal macrophages were then washed with 1×PBS (Ca2+- and Mg2+-free) and plated with DMEM containing 10% FBS, 100 U/ml of penicillin, and 100 U/ml of streptomycin at 37℃ in a 5% CO2 incubator.
Preparation of microencapsulated OVA
OVA-containing microspheres were prepared by a solvent-evaporation method (11). Briefly, albumin from chicken egg white was dissolved in 3% polyvinyl alcohol (4 mg/ml), and poly (DL-lactide-co-glycolide) (PLGA; lactide : glycolide=50 : 50; Sigma-Aldrich, St.Louis, MO, USA) was dissolved in a mixture of acetone and ethanol (9 : 1) (5%). The concentration of OVA was determined using a micro-bicinchoninic acid assay kit (Pierce, Rockford, IL) according to the manufacturer's instructions after lysing the microspheres in a lysis buffer containing 0.1% SDS and 0.1 N NaOH. For the phagocytosis assay, microspheres containing both OVA and fluorescein isothiocyanate (FITC) were prepared by adding FITC (final, 5 mg/ml) to a mixture of acetone and ethanol (9 : 1) together with PLGA (final, 5%).
MHC class I-restricted presentation assay of exogenous OVA
DCs were cultured in the presence of different concentrations of CME for 2 hrs in 96-well plates (1×105 cells/ml), and then combined with OVA-microspheres (6.25µg/min as OVA). After 6 hrs incubation at 37℃, the plate was washed twice of pre-warmed PBS, and then fixed with ice-cold 1% paraformaldehyde for 5 min at room temperature. The plate was washed and CD8OVA1.3 cells (1×105 cells/ml) were added. After overnight incubation at 37℃, the plate was centrifuged at 1,800 rpm, and the culture supernatant was collected and assayed for IL-2 content using an IL-2 ELISA kit (eBioscience, San Jose, CA, USA).
MHC class II-restricted presentation assay of exogenous OVA
DCs and peritoneal macrophages were cultured in the presence of different concentration of CME for 2 hrs in 96-well plates (1×105 cells/ml), and then combined with OVA-microspheres (6.25µg/min as OVA). After 6 hrs incubation at 37℃, the plate was washed twice of pre-warmed PBS, and then fixed with ice-cold 1% paraformaldehyde for 5 min at room temperature. The plate was washed and DOBW cells (1×105 cells/ml) were added. After overnight incubation at 37℃, the plate was centrifuged at 1,800 rpm, and the culture supernatant was collected and assayed for IL-2 content using an IL-2 ELISA kit.
Phagocytosis assay
DCs were cultured in the presence of different concentrations of CME for 2 hrs in 6-well plates (2×106 cells/well), and then combined with microspheres (average diameter, 300 nm) containing both OVA and FITC. After 2 hrs, unphagocytozed microspheres were removed by washing with pre-warmed PBS. The plate was chilled on ice for 20 min, and subsequently, the cells remaining on the bottom of plate wells were harvested by rough pipetting with cold PBS, washed with 10 ml of cold PBS to remove the extra particular OVA, and then fixed with 1% paraformaldehyde. Flow cytometric analysis was performed on a FACS Canto flow cytometer (Beckman coulter, Brea, USA).
Phenotype analysis
DCs were cultured in the presence of different concentrations of CME for 2 hrs in 6-well plates (2×106 cells/well). The plate was chilled on ice for 20 min, and subsequently, the cells remaining on the bottom of plate wells were harvested by rough pipetting with cold PBS. The cells were stained with monoclonal antibodies recognizing murine cell surface molecules after blocking of FcR-binding anti-CD16/CD32 monoclonal antibody, and flow cytometry analysis was performed on a FACS Canto flow cytometer. The monoclonal antibodies, anti-H-2Kb, anti-I-Ab, and isotype-matched control antibodies were purchased from BD Biosciences (San Jose, USA).
Preparation of nuclear extracts
After cultured cells were collected and washed twice with cold PBS, they were resuspended in hypotonic buffer (10 mM HEPES, pH 7.9, 10 mM KCl, 1.5 mM MgCl2, 0.2 mM PMSF, 0.5 mM DTT, 10µg/ml aportinin). After 15 min incubation on ice, the cells were lysed by the addition of 0.1% NP-40 and vigorous vortexing for 1 min. The nuclei were pelleted by centrifugation at 12,000×g for 1 min at 4℃ and resuspended in high salt buffer (20 mM HEPES, pH 7.9, 25% glycerol, 400 mM KCl, 1.5 mM MgCl2, 0.2 mM EDTA, 0.5 mM DTT, 1 mM NaF, 1 mM sodium orthovanadate). The supernatant fluid was stored in aliquots at -70℃.
Western blot analysis
Peritoneal macrophages were washed with PBS and lysed by lysis buffer (1% SDS, 1.0 mM sodium vanadate, 10 mM Tris-Cl buffer, pH 7.4) for 5 min. 20µg of protein from the cell lysates was applied to 8~12% SDS-polyacrylamide gels and then transferred to nitrocellulose membranes. The membranes were blocked with 5% skim milk in PBST solution for 1 hr. They were then incubated with anti-IL-1β, anti-IL-6, anti-TNF-α, anti-i-NOS, anti-COX-2, and anti-NF-κB monoclonal antibodies (Santa Cruz Biotechnology Inc., Santa Cruz, California, USA) for 2 hrs and washed 3 times with PBST. After incubation with an alkaline phosphatase-labeled secondary antibody (Abcam, Cambridge, USA) for 2 hrs, the bands were visualized using a Western Blot Kit with an alkaline phosphatase substrate (Vector, Burlingame, USA).
Data analysis
Data were expressed as mean±standard deviation. Statistical significance between the groups was determined by a paired t-test and one-way ANOVA for repeated measures. Results of p<0.05 were considered statistically significant. Data were assessed using SPSS (version 15.0, SPSS Inc., Chicago, Illinois).
RESULTS
CME enhanced MHC-restricted presentation of exogenous OVA
The effects of CME on MHC class I-restricted presentation of exogenous OVA in DC2.4 cells and BM-DCs were examined. DCs were treated with CME for 2 hrs, after which micro-encapsulated OVA were added into cultures for phagocytosis to examine whether exogenous antigen presentation pathways could be affected by CME. The cells were washed and then an MHC class I-restricted presentation assay was performed with CD8OVA1.3 cells. After 18 hrs incubation at 37℃ and 5% CO2, the supernatant was collected, and then assayed for IL-2 content using an IL-2 ELISA kit. As shown in Fig. 1, CME enhanced MHC class I-restricted OVA presentation. The effects of CME on the class II presentation of exogenous OVA in both BM-DCs and peritoneal macrophages were examined. DCs were generated from bone marrow cells of BALB/C mice and peritoneal macrophages were elicited by injection of thioglycolate into mouse peritoneum. Subsequently cells were treated for 2 hrs with CME, allowed to phagocytose OVA-microspheres for 6 hrs, and then unphagocytosed OVA microspheres and CME were removed by washing, after which a class II MHC-restricted presentation assay was performed with DOBW cells. After 18 hrs incubation at 37℃ and 5% CO2, the supernatant was collected, and then assayed for IL-2 content using an IL-2 ELISA kit. As shown in Fig. 2, MHC class II-restricted presentation of exogenous OVA was significantly increased by CME in both BM-DCs and peritoneal macrophages.
CME did not change the phagocytic activity of DCs
To prove that the augmentation of antigen presentation by CME was not due to increased phagocytic activity of DCs, FITC-conjugated OVA microspheres were prepared according to the method described above. DCs were incubated with CME for 2 hrs. Then microspheres containing both OVA and FITC were added. After 2 h, DCs were washed and then harvested. The level of phagocytosis was determined by a flow cytometry. As shown in Fig. 3, flow cytometric analysis of the harvested cells neither of the drugs induced the changes in the phagocytic activity of DCs.
CME enhanced the expression of MHC molecules
To measure the effects on the expression level of total MHC molecules of DCs which were pre-treated with the indicated concentration of CME, DCs were stained with anti-H-2Kb (MHC I molecule) and anti-I-Ab (MHC II molecule). As shown in Fig. 4, CME increased the expression of MHC class I and II molecules.
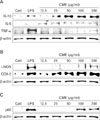
CME increased pro-inflammatory cytokines and cytokine related enzymes via NF-κB activation
To further investigate the important role of CME on the immune response, inducing peritoneal macrophages for overnight with LPS or CME resulted in the production of proinflammatory cytokines. As shown in Fig. 5, CME concentration-dependently induced the expression of IL-1β, IL-6, and TNF-α protein (Fig. 5A) as well as cytokine-related enzymes such as, iNOS and COX-2 protein (Fig. 5B). Also, to investigate whether CME could affect nuclear translocation of NF-κB, a western blot analysis for NF-κB p65 was carried out with cell lysate in macrophages (Fig. 5C). The amount of NF-κB p65 markedly increased upon exposure to LPS alone; CME also increased NF-κB p65.
DISCUSSION
Cordyceps militarys is a herbal ingredients, which has been used as a medicinal product for a long time in Oriental medicine for patients suffering from cancer (12) Some immunomodulatory and antitumor activities of CM have been established (13,14). Also, CM has been shown to inhibit tube formation of endothelial cells in vitro and in vivo and to repress the growth of melanoma cells in mice (12). However, little is known about its immunomodulatory effects or the mechanism of its effects on DCs.
This is the first report of which we are aware concerning the effects of CME on the both DCs and peritoneal macrophages. In this study, we have shown that DCs exposed to CME are enhanced with respect to MHC-restricted exogenous antigen presenting capability. Further, CME increased MHC class I and class II-restricted exogenous antigen presenting capability of DCs leading to induced expression of total MHC molecules. However, CME did not affect phagocytic activity of exogenous antigen.
Since T cells can only recognize antigens presented on MHC molecules, the impact of the inhibition of MHC-restricted antigen presentation must be far-reaching. DCs, a professional APCs, play key role in the initiation of primary immune responses (7) and are capable of processing cell-associated antigens for presentation on MHC class I molecules (15). In vitro, several forms of antigens can access the exogenous pathway for cross presentation, apparently through phagocytic or nonphagocytic mechanisms (16). Further, DCs can acquire and process antigens in the periphery and migrate to secondary lymphoid tissues where they prime primary T cell responses. The classical paradigm of antigen presentation by APCs is that endogenous antigens are presented via MHC class I molecules to CD8 T cells, whereas exogenous antigens are presented via CD4 T cells (17). However, APCs, process exogenous antigens for presentation by MHC class I molecules to CD8 T cells. This process, termed cross presentation, may be a mechanism by which naïve T cells can be primed to antigens that are present in nonprofessional APCs (18,19). The results presented in this paper demonstrate that short-term exposure of DCs to CME significantly enhances both MHC class I and II-restricted antigen presentation capabilities of DCs. Based on the importance of DCs in the initiation of T cell responses and the current finding, which demonstrated that short-term exposure to CME induced MHC-restricted antigen presentation, we are tempted to speculate that the therapeutic efficacy of CME is due, at least in part, to the enhancement of the antigen presenting capability of DCs.
Also, macrophages are unique cells in the immune system in that they not only can initiate immune responses, but also can serve as effector cells. Furthermore, activated macrophages become more efficient antigen presenting cells because they express increased levels of class II MHC molecules and co-stimulatory molecules (6,20,21).
Thus, based on the results of the present study, which showed that CME can regulate MHC class I and class II-restricted presentation of exogenous antigens through an increase in the expression of proinflammatory cytokine proteins, but do not affect phagocytosis in DCs, we are tempted to speculate that some of therapeutic efficacies such as immune-enhancer agents for immunodeficiency diseases.




 XML Download
XML Download