

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Granulocyte-colony stimulating factor (G-CSF) is extensively used to improve neutrophil count during anticancer chemotherapy [1-3]. In addition to the well-known effect of hematopoietic stem cell mobilization, the roles of G-CSF are diverse: activation of cellular function, inhibition of apoptosis, and increase in cell adhesion [4, 5]. We previously reported increased expression of G-CSFR in AML1/ETO+ AML cells, suggesting that cells with G-CSFR expression may proliferate easily in response to therapeutic G-CSF [2]. In the present study, we investigated the effects of the most commonly used commercial forms of G-CSF (glycosylated lenograstim and nonglycosylated filgrastim) on various leukemic cell lines. Moreover, we screened for the expression of G-CSFR mRNA in 38 solid tumor cell lines using real-time PCR because solid tumors of non-hematopoietic origin have been shown to express G-CSF or G-CSFR [6-13]. Detecting G-CSFR expression in primary tumor cells would allow careful monitoring of patient condition and aid in early detection of proliferation and/or differentiation. To our knowledge, this is the first study providing a quantitative comparison of G-CSFR mRNA expression in various solid tumor cells.
MATERIALS AND METHODS
1. Cell lines
Kasumi or CTV-1 cells, acute myeloid leukemia (AML) cells with or without the AML1/ETO gene rearrangement respectively; K562 chronic myelogenous leukemia (CML) cells; and the U266 multiple myeloma (MM) cell line were obtained from the Korean Cell Line Bank (KCLB, Seoul, Korea) and the America Type Culture Collection (ATCC, Rockville, MD). The cDNA from various solid tumor cell lines including SNU-201, U87MG, and A172, glioblastomas; Hs683, brain glioma; IMR-32, neuroblastoma; A375P, MDA-MB-435, and Malme-3M, melanomas; A498 and 293, renal cell carcinomas; AGS, gastric adenocarcinoma; DU-145, prostate carcinoma; FaDu and SNU-1041, squamous cell carcinomas of the pharynx; HCC-95 and SK-MES-1, squamous cell carcinomas of the lung; HeLa, adenocarcinoma of the cervix; Hep-2, epidermoid carcinoma of the larynx; SNU-899, squamous cell carcinoma of the larynx; HepG2, hepatoblastoma; SNU-368, 423, 449, and 878, hepatocellular carcinomas; OVCAR-3, ovarian adenocarcinoma. SNU-119, ovarian cystadenoma; RPMI2650, nasal squamous cell carcinoma; RT4, transitional cell carcinoma; SNU-410, pancreatic carcinoma; SNU-175 and C2B, carcinoma of colon; UV2237M, fibrosarcoma; MCF-7, MCF-10A, BT20, MDA-MB-231, HCC1954 and T47D, breast carcinomas were kindly provided by the Korean Cell Line Bank (KCLB, Seoul, Korea).
2. Real-time quantitative PCR measurement of G-CSFR
Real-time quantitative PCR was performed using a Universal TaqMan Probe Master Mix (Applied Biosystems Foster City, CA, USA). Amplification was performed at 50℃ for 2 min and 95℃ for 10 min, followed by 40 cycles at 95℃ for 30 sec, 60℃ for 30 sec, and 72℃ for 30 sec. TaqMan analysis was used to detect CSF3R (Hs00167918_m1) and GAPDH (Hs99999905_m1) mRNA expression using primers and conditions designed by assays-on-demand gene expression products (Applied Biosystems, USA). Each of the 384-well real-time quantitative PCR plates contained serial dilutions (1, 1/2, 1/4, 1/8, and 1/16) of cDNA, which were used to generate relative standard curves for CSF3R and GAPDH. The G-CSFR expression was normalized to GAPDH expression. The real-time PCR analysis was performed using an Applied Biosystems Prism 7900 Sequence Detection System (Applied Biosystems, USA). Data were analyzed using ABI Prism 7700 SDS software (version 1.0). The levels of G-CSFR expression were confirmed in 3 independent experiments.
3. Cell proliferation assay
The proliferation of cells was evaluated using a Cell-Titer 96® Non-Radioactive Cell Proliferation Assay (Promega Co., Madison, WI, USA) according to the manufacturer's protocol. Briefly, the cells were suspended to obtain a final concentration of 1×105 cells/mL, and 500 µL of this suspension was incubated at 37℃ for 48-72 h in a humidified, 5% CO2 atmosphere. After 4 h of incubation in a dye solution, 100 µL of solubilization solution/stop mix was added, and the absorbance was recorded at a wavelength of 570 nm. Analysis of cell proliferation using an EdU assay was also performed. A Click-iT™ EdU Alexa Fluor Flow Cytometry Kit (Invitrogen, Eugene, OR, USA) was used in accordance with the manufacturer's instructions. Briefly, G-CSF-treated or untreated Kasumi-1 and CTV-1 cells were incubated with 10 µM EdU in culture media at 37℃ for 60 min. The cells were harvested, fixed, and permeabilized with 5% Triton X-100 for 30 min, and then stained with Alexa Fluor 647 dye in the dark for 30 min. Fluorescence intensity was measured by flow cytometry (BD Biosciences, San Jose, CA), and the percentage of cell proliferation was determined using FlowJo flow cytometry analysis software (Tree Star Inc., Ashland, OR, USA). The results were validated with 2 repeated experiments.
4. Differentiation study of granulocytic series by flow cytometry
Cell suspensions with the same cell density were placed in sterile culture dishes and treated with 2 forms of G-CSF (filgrastim, lenograstim) at concentrations of 0, 10, 50, and 100 ng/mL for 2 weeks. At 0, 3, 7, and 14 d after G-CSF treatment, cells were harvested and analyzed by triple-staining with fluorescein isothiocyanate, phycoerythrin, and PerCP-conjugated monoclonal antibodies for CD11b and CD66b (Becton Dickinson Biosciences, San Diego, CA, USA and DakoCytomation, Glostrup, Denmark). Negative controls included a mouse isotype-matched non-relevant immunoglobulin. The samples were analyzed by flow cytometry (FACSCanto, Becton Dickinson, Franklin Lakes, NJ, USA). The results were validated by 2 repeated experimentation.
RESULTS
1. Expression of G-CSFR in leukemic and solid tumor cell lines
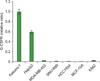
We analyzed G-CSFR expression in Kasumi-1 (AML with AML1/ETO gene rearrangement), CTV-1 (AML without AML1/ETO gene rearrangement), K562 (CML), and U266 cell lines (MM). Kasumi-1 and K562 cells expressed G-CSFR mRNA whereas CTV-1 and U266 did not. Compared to G-CSFR expression in Kasumi-1 cells, K562 expressed a relatively small amount of G-CSFR mRNA (relative expression: 0.02). Among the 38 solid tumor cell lines tested, 5 (13.1%) (hepatoblastoma [HepG2], melanoma [MDA-MB-435], squamous cell carcinoma of larynx [SNU-899], and breast cancer [HCC 1954 and MCF-10A] cell lines) expressed G-CSFR mRNA, with relative G-CSFR expressions of 0.76, 0.03, 0.01, 0.01, and 0.002, respectively. Expression of G-CSFR in the HepG2 cell line was high and comparable to that in the Kasumi-1 cell line (Fig. 1).
2. G-CSF-induced proliferation of leukemic cells
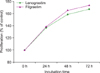
Both forms of G-CSF (filgrastim and lenograstim) significantly and comparably stimulated the proliferation of AML1/ETO+ Kasumi-1 cells at increasing concentrations (10, 50, and 100 ng/mL) (Fig. 2). In K562 cells, treatment with G-CSF only resulted in a slight increase in proliferation at the highest dose of filgrastim. CTV-1 cells showed very mild proliferation, but only after treatment with lenograstim for 72 h at high concentrations (50 and 100 ng/mL). U266 cells showed no proliferation upon treatment with G-CSF (Fig. 2).
The EdU assay confirmed that, when compared to unstimulated controls, lenograstim and filgrastim (both at 10 ng/mL) increased the proliferation of Kasumi-1 cells (from 38.7% to 51.9% and 51.5%, respectively), whereas CTV-1 cells did not respond to treatment. Both filgrastim and lenograstim significantly stimulated the proliferation of AML1/ETO+ Kasumi-1 cells in a time-dependent manner (Fig. 3).
3. G-CSF-induced differentiation of leukemic cells
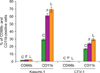
The differentiation effect was determined by analysis of mature granulocyte phenotype marker expression (CD11b and CD66b) by flow cytometry. G-CSF treatment doubled the proportion of CD11b-positive cells in Kasumi-1 cells, whereas the proportion of CD11b-positive cells only mildly increased in CTV-1 cells after 14 d of incubation with G-CSF at 100 ng/mL. Although CD11b-positive cells also increased in unstimulated control cells (29%), the increase in CD11b-positive cells was much more prominent in Kasumi-1 cells after filgrastim and lenograstim treatment (61% and 69%, respectively) (Fig. 4). Expression of CD66b was not significantly affected in either cell line regardless of the concentration of G-CSF, type of G-CSF, or incubation time (Fig. 4).
DISCUSSION
Many growth factors play pivotal roles in cell proliferation, migration, and differentiation [14]. Although a possible stimulating influence on leukemic cells remains questionable, most studies have reported that G-CSF is a safe agent that improves neutrophil count, thereby reducing the incidence of documented infection without regrowth of leukemic cells or other negative effects [15-18]. In the present study, we evaluated the effects of 2 forms of recombinant human G-CSF (rhG-CSF) available for clinical use: filgrastim is derived from Escherichia coli and has a non-glycosylated form, whereas lenograstim is derived from Chinese hamster ovary (CHO) cells and is glycosylated. The peak serum concentrations of G-CSF after administration of a standard dose of G-CSF (5 µg/kg) is found to range from 15 to 30 ng/mL [19, 20]; therefore, we used a range of G-CSF concentrations spanning this concentration (0, 10, 50, and 100 ng/mL). The proliferation effect of G-CSF was prominent in Kasumi-1 cells, and the 2 forms of G-CSF showed similar effects. This might be due to the high expression of G-CSFR in AML1/ETO+ Kasumi-1 cells, as reported in previous studies [2, 5]. Meanwhile, the proliferation was not stimulated in CTV-1 cells or U266 cells, which did not express G-CSFR. K562 cells with low-level expression of G-CSFR mRNA showed mild proliferation only at 100 ng/mL filgrastim after 72 h; however, this concentration cannot be applied in a clinical setting and is also much higher than the estimated serum concentrations in patients after G-CSF administration.
In addition to a proliferative effect, we noted that G-CSF induced differentiation in AML1/ETO-positive cells with a high level of G-CSFR expression. There have been several reports on the differentiation effect of G-CSF, evidenced by morphologic changes and immunophenotypic changes [21-25]. In the present study, although mild changes in CD11b expression were observed in unstimulated control cells and in stimulated CTV-1 cells, there was a prominent increase of CD11b expression in G-CSF-treated Kasumi-1 cells. However, the expression of CD66b was not affected. During the normal process of differentiation of neutrophils in bone marrow, CD66b is expressed from CD34-myeloblasts, reaching its highest level of expression at the myelocyte stage and decreasing thereafter [26], whereas CD11b is expressed at a later stage and its expression increases during maturation [27]. Thus, the differentiation pattern found in the present study (CD11b+/CD66b-) would not be seen in a normal process and suggests that the in vitro differentiation induced by G-CSF was abnormal and incomplete. Induction of CD11b expression by G-CSF has been previously reported [22, 23]. Given that not all patients with AML1/ETO+ AML show prominent differentiation in response to exogenous G-CSF, it is inferred that other factors such as microenvironments have to be taken into consideration for differentiation in vivo.
Many studies have demonstrated the expression of G-CSFR in tumor cells or autocrine secretion of G-CSF in non-hematopoietic tumors such as colon cancer, ovarian cancer, squamous cell cancer, malignant melanoma, and sarcoma [6-13, 28-30]. In these reports, G-CSF was shown to stimulate proliferation and angiogenesis, and subsequently enhance malignant potential [6, 13]. Owing to the potential risk of stimulation of proliferation by G-CSF, information concerning G-CSFR expression in tumor cells would be helpful in the management of cancer patients. Here, we performed quantitative G-CSFR mRNA expression analyses in various solid tumor cell lines. Among the solid tumors, 13.1% expressed G-CSFR. Of note, G-CSFR expression in the hepatoblastoma cell line HepG2 was high and comparable to that in the AML1/ETO+ Kasumi-1 cell line [2, 5]. However, such expression, especially when low, should be confirmed through additional testing and the statistical relevance of low expression needs to be validated.
In conclusion, G-CSFR expression and the proliferative effects of G-CSF on various malignant cells were demonstrated in the present study. Therefore, G-CSF should be used with caution in patients with hematopoietic or non-hematopoietic tumors with high G-CSFR expression. Accordingly, we suggest that screening for G-CSFR before administering G-CSF would be helpful in minimizing the risk of tumor proliferation. Expression levels of G-CSFR in primary tumor tissues should be evaluated by further study.

 XML Download
XML Download