

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Graft-versus-host disease (GVHD) is a leading cause of mortality and morbidity following allogeneic hematopoietic cell transplantation (HCT). It is caused by mature donor T lymphocytes, which recognize the tissues of the recipient as foreign, causing a severe inflammatory disease most often characterized by rash, diarrhea, and liver disease. GVHD is classified into acute or chronic forms by its symptoms, the amount of damage it has caused to the liver, skin and mucosa and gastrointestinal tract as well as its timing. While acute GVHD usually refers to disease presenting within the first 100 days post-transplant, chronic GVHD symptoms may occasionally present as early as 50-60 days post-transplant and symptoms may overlap, typified by broader tissue damages often involving connective tissues and exocrine glands.
GVHD is particularly virulent when there is a mismatch of major histocompatibility complex (MHC) class I or MHC class II antigen between the donor and the host. MHC molecules are highly polymorphic glycoproteins that present peptide antigens to T cells. While MHC class I molecules present peptides generated in the cytosol to CD8 T cells, MHC class II molecules present peptides degraded in the intracellular vesicles to CD4 T cells. Thus, it is commonly recommended that HCT be undertaken, when there is a human leukocyte antigen (HLA) (MHC of human) match between the donor and the host. However, GVHD can also occur in the context of disparities between minor histocompatibility antigens, which are peptides of polymorphic cellular proteins bound to MHC molecules that can lead to graft rejection when they are recognized by T cells. One set of proteins that induce minor histocompatibility responses is encoded on the male-specific Y chromosome and the responses induced by these proteins are called H-Y. Female anti-male minor histocompatibility response can occur in female transplant recipients, since Y chromosome-specific genes are not expressed in females.
Because GVHD is characterized by tissue damage caused by T-cell mediated immune attack, immunosuppression is commonly used in the setting of HCT. The use of immunosuppressive agents is to reduce the severity of GVHD with the potential to induce donor-specific tolerance and avoid increased risk of infection and cancer in transplant recipient. Donor T cell activation requires two signaling events including binding of the T-cell receptor (TCR) to an allogeneic peptide presented by the host MHC as well as co-stimulatory signals delivered by antigen presenting cells (APCs). Co-stimulatory signals involve the interaction of CD28 on the T cell with CD80 or CD86 on the APCs. CD28 activates a signal transduction pathway, acting through phosphoinositide 3-kinase (PI3K) to provide its co-stimulatory signal for T cell activation. Dendritic cells or macrophages are major hematopoietic originated APCs and a capacity of these cells to activate antigen presentation is determined by the innate immune mechanisms that sense various types of microbial ligands or endogenous danger signals. With the discovery of Toll-like receptors (TLRs) and nucleotide binding oligomerization domain (NOD)-like receptors (NLRs), the role of innate immunity in the regulation of adaptive immunity and modulation of immune tolerance has become an intense area of current investigation in the field of immunology.
TLRs have been extensively studied for their ability to activate signaling pathways in response to microbial or viral infection and to provide a link between innate and adaptive immunity, as TLR signals augment antigen presentation by innate immune cells [1]. Following ligand recognition or cellular disruption, these receptors activate downstream signaling pathways, such as nuclear factor kappa B (NF-κB), mitogen activated protein kinase (MAPK) and type I interferon (IFN) pathways, which result in the up-regulation of pro-inflammatory cytokines and chemokines that are important in inflammatory and antimicrobial responses. In addition to TLRs, NLRs also function to recognize pathogens and form a multi-protein intracellular complex with caspase-1, known as inflammasome, which lead to release of interleukin-1β (IL-1β), interleukin-18 (IL-18) and interleukin-33 (IL-33) [2].
Exploration of the potential role of TLRs/NLRs in the outcome of GVHD is in its early stages. In addition to their role in host defense against pathogenic microorganisms, TLRs/NLRs also sense endogenous ligands/danger signals [3] and this danger sensing may play a major role in survival and maintenance of the grafted organs [4]. Furthermore, studies of gastrointestinal GVHD show that the loss of intestinal homeostasis or tissue integrity contributed significantly to increased systemic severity of GVHD and this could be mediated by TLR4 [5] or NOD2 [6]. Given that polymorphisms in TLRs/NLRs are associated with increased risk of severe GVHD, understanding the mechanisms by which TLRs/NLRs play a role in GVHD pathophysiology is essential.
Recent advances in our understanding of how innate immune receptors recognize pathogen-associated molecular patterns (PAMPs) as well as danger-associated molecular patterns (DAMPs) and trigger adaptive immune response may provide a new insight into molecular mechanisms by which a specific modulation of these pathways can be used as novel therapeutic approaches for GVHD. We discuss our current knowledge of the biology of the TLRs/NLRs and the mechanisms of their pathogen recognition as well as non-pathogen recognition function. Furthermore, we focus on their potential role in pathophysiology of GVHD and discuss future therapeutic targets.
Go to : 
INTRODUCTION TO TOLL-LIKE RECEPTORS (TLRs)
Toll-like receptors (TLRs) are innate immune receptors capable of sensing various components of microbial organisms, such as lipid, carbohydrates, peptides, and nucleic acid. As evolutionarily conserved molecules, TLRs were first described in vertebrates as homologous proteins to insect molecule, Toll that stimulates the secretion of antimicrobial peptides in Drosophila Melagaster [7]. To date, 11 members of TLRs have been identified in human (13 TLRs in mouse) and their expression patterns are quite diverse in that they are expressed on both innate and adaptive immune cells [1]. While some TLRs are expressed extracellularly, expression of other TLRs (such as 3,7/8,9) are limited to endocytic or intracellular compartments. Most TLRs have transmembrane domains, composed of N-terminal extracellular leucine-rich repeats (LRRs) that are responsible for recognition of specific pathogen components, a membrane-spanning domain and a C-terminal cytoplasmic domain similar to the cytoplasmic region of the interleukin-1 (IL-1) receptor, known as the Toll/IL-1 receptor (TIR) domain, which is required for downstream signaling. TLRs (except for TLR3), as well as IL-1R and IL-18R family members utilize MyD88 as an adaptor molecule for downstream signaling [1].
TLRs recognize diverse microbial patterns and have broad specificity, detecting many related molecular structures. TLR2 can either dimerize with TLR1 or TLR6. While TLR2/6 heterodimers recognize diacyl lipoproteins from Mycoplasma species [8], TLR1/2 heterodimers recognize triacyl lipoproteins from various bacteria, including B. burgdorferi [9]. TLR3, which is found in endosomal compartments, recognizes double-stranded RNA (dsRNA) [10]. TLR4 mainly recognizes lipopolysaccharides (LPS) of gram-negative bacteria [11] and interacts with CD14 and MD2 [12, 13]. TLR5 specifically recognizes bacterial flagellin [14]. Both TLR7 and 8 sense single-stranded RNA (ssRNA) [15], whereas TLR9 is responsible for the recognition of unmethylated CpG nucleotides [16, 17]. TLR11 detects profilin and plays an important role in host defense against uropathogenic bacterial infection [18, 19].
TLR signaling is generally divided into two pathways (Fig. 1); MyD88-dependent and TRIF-dependent. Except for TLR3, TLRs utilize MyD88 as an adaptor molecule for signaling, leading to NF-κB-dependent cytokine production. On the other hand, TLR3 (and TLR4) uses a MyD88-independent signaling pathway that involves the adaptor molecule called TIR-containing adaptor inducing IFN-β (TRIF), which can either promote the activation of NF-κB pathway or the induction of type I interferon (IFN-α/β) [1]. Signaling pathways downstream of TLRs are shown in Fig. 1. In the case of MyD88-dependent signaling pathways, TIR domains of TLRs interact with MyD88 that recruits members of the IL-1R-associated kinase (IRAK) family. IRAK activation results in targeting downstream TNF receptor-associated factor 6 (TRAF6), through the recruitment of transforming growth factor β activated kinase 1 (TAK1) and TAK1-binding protein 2 (TAB2). These molecules ultimately lead to activation of the upstream kinases for mitogen-activated protein kinases (MAPK) and activation of NF-κB occurs, when IκB kinases (IKK) complex phosphorylates IκB, which leads to nuclear translocation of NF-κB.
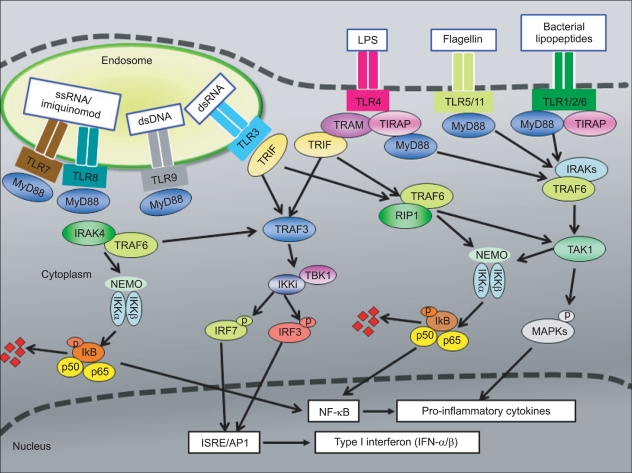 | Fig. 1Toll-like receptor (TLR) signaling pathway. Toll-like receptors (TLRs), except for TLR3, use a MyD88-dependent signaling pathway to induce NF-κB activation. TLR3, (and TLR4), on the other hand, can activate MyD88-independent, TRIF-dependent pathway to induce NF-κB or type I interferon (IFN) activation. MyD88 recruits TRAF6 and members of the IRAK family, leading to the activation of the TAK1 complex. The activated TAK1 complex then activates the IKK complex, consisting of IKKα, IKKβ and NEMO (IKK-γ), which catalyze the phosphorylation of IκB proteins. IκBs are degraded by the proteasome, allowing NF-κB to translocate into the nucleus. Simultaneously, the TAK1 complex activates the MAPK pathway, which results in the phosphorylation and activation of AP-1. NF-κB and AP-1 control inflammatory responses through the induction of inflammatory cytokines. The signaling cascade triggered by TLR3 uses MyD88-independent, TRIF-dependent pathway. TRIF recruits TRAF3, which then interacts with TBK1 and IKKi. These kinases mediate phosphorylation of IRF3. Phosphorylated IRF3 dimerizes and translocates into the nucleus to regulate transcription. TRIF also interacts with TRAF6 and RIP1, which mediate NF-κB activation. Activation of the IRF3, NF-κB and MAPK pathways results in the induction of type I IFN, particularly IFN-β.
|
Similar to MyD88-dependent pathway, TRIF-dependent pathway also activates NF-κB. In addition to NF-κB activation, the TRIF-dependent pathway can also lead to the induction of type I interferon through phosphorylation and activation of the transcription factors, interferon regulatory factor 3 (IRF3) and IRF7 [20, 21]. TRIF is used by TLR3 or TLR4 and can directly bind to TRAF6 via its TRAF6-binding motifs in the N-terminal region and TRAF6 then activates TAK1 in a manner similar to that in the MyD88-dependent pathway [22, 23]. TRIF can also recruit a signaling complex involving the non-canonical IKKs, TBK1 and IKKi, which induce IRF3 activation, mediated by TRAF3 [24].
A crucial role for TLRs in regulating the immune response is implicated in many cellular processes and diseases. Signaling via TLRs can initiate inflammatory cascades in response to pathogens or cellular stress, and trigger the activation of adaptive immune responses by regulating expression of co-stimulatory molecules on APCs, such as CD40 or CD80/86, and thereby enhancing antigen presentation [25]. Recent findings indicate that TLRs are involved in the process of acute allograft rejection and that their activation can prevent transplantation tolerance. We will review the expression of TLRs and the impact of TLR signaling on grafted organs in studies using both animal experiments and human clinical genetic association results. We then discuss possible mechanisms by which TLRs are involved in the rejection phenomenon and lead to increased alloimmune responses during GVHD.
Go to :
THE ROLE OF TLRs IN GVHD
Efficient priming of adaptive immune response depends on antigen presentation and stimulation of co-stimulatory molecules. TLRs regulate the expression of these co-stimulatory molecules and subsequently control the adaptive immune response, thereby influencing T cell activities [25]. Thus, because of TLRs' role in the control of adaptive immune response, it was hypothesized that TLR signals may influence the activation of donor T lymphocytes and thus exacerbate the outcome of GVHD. Moreover, TLRs have been shown to be involved in regulatory mechanisms for ischemia-reperfusion injury (IRI), which is a major cause of delayed allograft function in solid organ transplantation (SOT) and an important inducer of acute and chronic transplant rejection [26-30]. Evidences suggesting a role for TLRs in immune tolerance against allograft in SOT is based on studies from both animal experiments using knockout mice and human genetic single nucleotide polymorphism (SNP) analysis.
LPS is a TLR4 ligand and LPS-mediated TLR4 activation leads to pro-inflammatory cytokine release [1]. The finding that LPS signals could influence the outcome of GVHD facilitated the use of LPS antagonist at the time of transplantation to reduce the risk of GVHD [31]. The importance of TLR signaling in graft intolerance using animal transplantation model was confirmed when Goldstein et al. reported the tolerance of MyD88-deficient mice towards H-Y minor histocompatibility antigen-mismatched transplant, suggesting that the GVHD in H-Y minor histocompatibility antigen-mismatched transplant in mice is dependent on the presence of TLR signaling [32]. The authors further investigated whether the absence of acute allograft rejection from the MyD88-deficient recipients could be restored by the adoptive transfer of WT-activated spleen cells. WT cells could re-activate the alloimmune response, confirming that TLR signaling exacerbates the alloimmune tolerance. Thus, these results suggest that the defect in the MyD88-deficient recipients most likely arises in the initiation phase of the alloimmune response because there are reduced numbers of mature dendritic cells migrating into draining lymph nodes, resulting in MyD88-deficient mice's inability to generate alloreactive T cells and thus cause the intolerance. Furthermore, consistent with this study, even for MHC-mismatched SOT, TLR signaling was found to be important for dendritic cell function and TH1 cell response, which influence the outcome of transplantation [33].
In addition to animal studies described above, the implication that TLR signaling may influence the severity and incidence of acute GVHD is also shown by human genetic association studies [34, 35]. An association between TLR4 mutation and increased risk for GVHD was observed for patients who received organ grafts from human leukocyte antigen (HLA)-matched donor siblings, and increased risk for gram-negative bacteremia as a result of TLR4 mutation was also reported in these patients [34]. However, these associations are not statistically significant in recipients of HLA-matched sibling marrow transplants, prophylactically treated for infections and GVHD, suggesting that TLR4 polymorphisms may not cause more severe GVHD. However, another study indicates that there is a significant influence of mutations in TLR4 on the incidence and severity of acute GVHD in patients who underwent allogenic transplantation [35]. Interestingly, specific single nucleotide polymorphisms (SNP) in the coding region of the TLR4 gene in both the patient or donor side were associated with an increased risk for severe GVHD and intestinal GVHD. Based on the possibility that microbiota may contribute to GVHD outcome, the use of antibiotics during GVHD was evaluated and metronidazole and ciprofloxacin to decontaminate the intestine were found to be helpful for reducing the incidence of severe acute GVHD [35]. It is possible that differential composition of the intestinal microbiota from the recipients of HCT possibly contributes to the outcome of graft survival and maintenance. The finding that suggests a role for TLRs in the control of intestinal microbiota environment [36] could be also a contributing factor that influences the occurrence and severity of GVHD.
Besides TLR4, several other TLRs have been also implicated to play a role in GVHD pathophysiology. As mentioned earlier, TLR7/8 are known to recognize single-stranded RNA and induce anti-viral response. The expression of TLR7/8 is present on plasmacytoid dendritic cells (pDCs) [37], which function as major anti-viral APCs. Jasperson et al. show that pDCs are capable of antigen presentation in GVHD and express immunosuppressive enzyme, indoleamine 2,3-dioxygenase (IDO), that influences GVHD pathophysiology [38]. Interestingly, administration of TLR7/8 agonist induced IDO expression and reduced GVHD pathogenecity [39]. In addition, TLR9, a receptor that recognizes CpG DNA, was also shown to be involved in GVHD outcome. TLR9 ligation of recipient APCs by CpG DNA markedly accelerated GVHD lethality in a mouse transplantation model [40].
What are the mechanisms by which these TLR signals affect the outcome of GVHD? One possibility is that TLR-mediated activation of recipient APCs may lead to maturation and migration of APCs, thereby indirectly causing the activation of donor T lymphocytes. T cell activation is likely to be triggered by stimulation of TLR pathway, thereby promoting the maturation and migration of alloantigen-expressing DCs to draining lymphoid organs. Transplantation studies with MyD88-deficient or MyD88/TRIF-deficient mice supported the hypothesis that TLR function on APCs is important for T cell activity and GVHD outcome. Defective TLR signals from either MyD88-deficient or MyD88/TRIF-deficient mice led to prolonged graft survival and this was consistent with diminished migration of donor cells to draining lymph nodes and subsequently, with delayed infiltration of host T cells into the grafted tissue [32, 41].
Although previous studies mentioned above focused on TLR-mediated stimulatory effect on APCs that indirectly affect T cell response, there are studies to suggest direct effects of TLRs on T cell functions. Given that the expression of TLRs is not limited to innate immune cells, but is extended in different T cell subsets, including conventional αβ T cells, regulatory T cells, and γδ T cells as well as natural killer T cells, we can speculate that TLRs may be significant for induction of effector and regulatory function of T cells [42]. Stimulation of TLR ligand with purified CD4 T cells can directly promote activated CD4 T cell survival, suggesting that TLRs on T cells can directly modulate adaptive immune response [43, 44]. Furthermore, TLRs can act as a co-stimulatory receptor via T cell receptor (TCR) engagement and can enhance the priming or survival of alloreactive T cells [45]. Another evidence that TLR stimulation provides a signal for T cell activation comes from the study using the administration of the TLR9 agonist CpG or of the TLR2 ligand Pam3CSK4 at the time of skin graft transplantation. This co-administration resulted in increased production of interferon (IFN)-γ by alloantigen-stimulated splenocytes and impaired intra-graft regulatory T cells (Treg) recruitment, suggesting that innate immune signals prevented alloimmune tolerance against the graft [46].
In addition to TLRs' indirect or direct role in activation of T lymphocytes, TLRs may also participate in the control of intestinal microbiota, thereby contributing to the GVHD pathophysiology. Intestinal microbiota and endotoxin were proposed to have a significant impact on the APC activation and this could be a crucial and initiating step in the induction of alloreactions [47-49]. Compared with mice who died from acute GVHD, grown under conventional conditions early after transplantation, germ-free mice were resistant to developing acute GVHD, suggesting that intestinal microbiota could determine the susceptibility for GVHD severity [50]. Recent findings suggest that there is a significant association of TLR-mediated control of microbiota and the outcome of GVHD [47]. The beneficial role of the diversity of the intestinal microbiota on the outcome of GVHD has been suggested by Gerbitz et al. in that alteration of the intestinal microbiota may play an important role in the initiation of acute GVHD and the use of probiotics can ameliorate the outcome of GVHD [49]. Understanding the mechanisms by which the interaction of TLR-mediated innate immune activation with intestinal microbiota may shape the outcome of GVHD remains to be investigated in the future.
In summary, both animal transplantation model and human clinical genetic studies show that TLR-mediated signaling is likely to play an important role in the pathogenesis GVHD. TLRs induce antigen presentation, together with TLR-mediated expression of co-stimulatory molecules and inflammatory cytokines, and instruct development of antigen specific adaptive immunity, especially helper T cells. Therefore, alloimmune T cell responses can be enhanced by microbial and/or endogenous TLR ligands induced by IRI and tissue stress/injury at the time of transplantation. Elucidation of the factors controlling TLR-mediated events after transplantation could ultimately lead to novel therapeutic strategies that target TLR signals to reduce the severity of GVHD.
Go to :
INTRODUCTION TO NLRs
Together with TLRs, NLRs represent central platforms of innate immunity that link sensing of microbial pathogens and metabolic stress to the activation of pro-inflammatory cytokines, such as IL-1β, IL-18 and IL-33. NLR signaling results in the formation of large molecular scaffold complexes (called inflammasome) [51], which are intricately linked with inflammation/autoimmunity [52, 53] and crosstalk with TLR-mediated signaling events [54]. Mutations/polymorphisms in NLRs have been linked to auto-inflammatory or autoimmune diseases [52, 55] and thus this suggest that NLRs function beyond the recognition of pathogens and act as central components of immune system. Since the discovery of inflammasome [51], NLRs-related studies have been focused on identifying molecules that are components of inflammasome and understanding how these molecules recognizes danger signals. Activation of caspase-1 through autoproteolytic maturation is thought to be a mechanism by which the processing and secretion of IL-1β occurs [51], however, further studies are required to determine the relative contribution of each of inflammasome mechanisms to innate and adaptive immune responses in response to danger signals.
More than 20 members of NLR family have been identified in mammals so far [56], however, the ligands and functions of many of these receptors remain undefined. In addition to TLRs, NLRs function as intracellular sensors that recognize pathogen-associated molecular patterns (PAMPs) as well as danger-associated molecular patterns (DAMPs) including cellular stress, and tissue damage/injury. NLRs are characterized by C-terminal leucine rich domain, an intermediate NOD domain and an N-terminal effector region, comprising a protein-protein interaction domain, such as the caspase activation recruitment domain (CARD), pyrin (PYD) or baculovirus inhibitor of apoptosis repeat domain (BIR) domain. NLRs have been grouped into several subfamilies on the basis of the effector domains: NOD, NALPs, and IPAF/NAIPs. NODs and IPAF contain CARD effector domains, whereas NALPs and NAIPs contain pyrin (PYD) effector domains and three BIR domains, respectively [56].
NOD1/2 were first NLRs to be identified and characterized. NOD2 was first described as a Crohn's disease susceptibility gene [57, 58] and later, NOD1/2 have been shown to be important for anti-microbial host response against bacterial pathogens and they are known to recognize peptidoglycans (PGN), cell wall components of bacteria. NOD1 is a sensor for D-γ-glutamyl-meso-DAP dipeptide (iE-DAP), which is found in PGN of all Gram-negative and certain Gram-positive bacteria [59], whereas NOD2 recognizes the muramyl dipeptide (MDP) [60]. Loss of NOD1 caused increased susceptibility to Helicobacter pylori [61], while NOD2 deficient mice were more susceptible to Listeria monocytogenes infection [62, 63]. NOD2 is also regarded as a pivotal sensor molecule of the intestinal barrier, contributing to maintenance of intestinal homeostasis [64]. Once activated, NOD1 and NOD2 oligomerize and recruit the NF-κB activating kinase RIP2 through homotypic CARD-CARD interactions involving their N-terminal CARD motifs. RIP2 interacts with the regulatory NF-κB subunit, NEMO/IKK-γ, triggering IκB phosphorylation and NF-κB activation [62, 65] (Fig. 2).
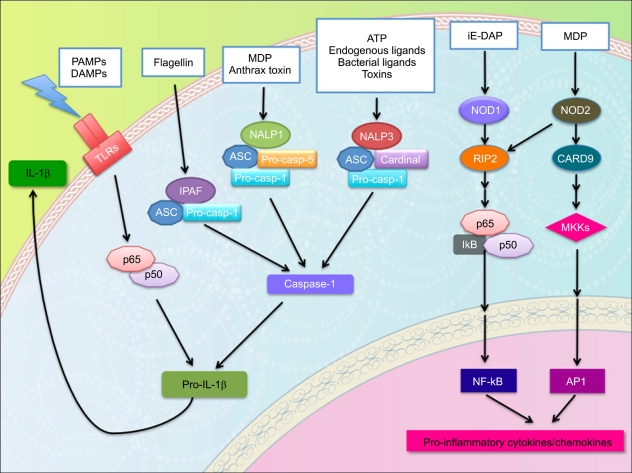 | Fig. 2NOD-like receptor (NLR) family and inflammasome. NOD1/2 sense peptidoglycan fragments (iE-DAP and MDP) respectively and activate signaling pathways via the serine/threonine RIP2 kinase through CARD-CARD homophilic interactions. Once activated, RIP2 mediates the ubiquitination of NEMO/IKKγ leading to the activation of NF-κB and the production of inflammatory cytokines. In addition to the NF-κB pathway, NOD2 stimulation induces the activation of MAPKs. Several NALPs, NALP1, NALP3, and IPAF, have been identified to sense various ligands and form a cytoplasmic mutiprotein complex called inflammasome. These NALPs recruit the adaptor protein ASC through their PYD domain, which in turn interacts with caspase-1 via a CARD-CARD interaction to form inflammasome. Consequently, inflammasome activation triggers the processing of pro-caspase-1 into a mature form, caspase-1 and active caspase-1 induces cell death and the processing of pro-inflammatory cytokine pro-IL-1β into IL-1β.
|
NALPs (NLRPs) are characterized by the presence of PYD effector domains and so far, include 14 members. The function of many NLRPs still remains unknown, however, several NALPs (NLRP1, NLRP3, and NLRC4) have been shown to participate in the formation of inflammasome. NALP1 (NLRP1) was shown to form cytoplasmic multicomplex with the adaptor called, apoptosis-associated speck-like protein containing a caspase recruitment PYCARD (ASC) and activate caspase-5. NLRP1 can be activated by two molecules, muramyl dipeptide (MDP) and the anthrax toxin [66, 67]. NALP3 (NLRP3) is the best studied NLR member, which has been shown to mediate caspase-1 activation in response to a variety source of stimulus; bacterial components [68-74], endogenous danger signals released by damaged cells or tissues [75-77], and pore-forming toxins [78]. NLRP3 also interacts with ASC to activate caspase-1 and NLRP3 signaling requires two stimulus, a cell priming signal from transcriptionally active TLR, NLR, or cytokine receptor, prior to activation of NLRP3 with pore-forming toxins, ATP or various endogenous danger signals. Similar to TLR5, IPAF (NLRC4) has been shown to respond to bacterial flagellin, a main component of the bacterial flagellum, restricting the proliferation of intracellular bacteria such as Salmonella typhimurium, Shigella flexneri, and Legionella pneumophila [79-82]. These bacteria use type III secretion system (TTSS) to secrete flagellin into cytoplasmic compartment where IPAF senses flagellin. NLRC4 lacks a PYD domain, and it could activate pro-caspase-1 directly, however, ASC is still required for full activation to induce the cleavage and secretion of mature IL-1β [83].
Recent studies focused on highlighting the role of NLRs in broader control of adaptive immune response and various disease states [84]. Many NLRs have been identified and proposed to function as sensors for PAMPs as well as DAMPs, released through damaged mucosal barriers or skin. Specifically, DAMPs-stimulated NLR activation may participate in enhancing the severity of GVHD, similar to the role of TLRs, thus, it is important to understand potential contribution of NLRs to GVHD outcome. Below, we summarize the current understanding of the function of NLRs in GVHD pathophysiology.
Go to :
ROLE OF NLRs IN GVHD
Similar to TLRs, NLRs were also shown to mediate the immune response to IRI [85-87]. Therefore, NLRs were proposed to influence the outcome of GVHD. Previous studies indicate an important role for NOD2 in contributing to susceptibility to GVHD after HCT [6, 35, 88-91] and mutations in NLRP2 and NLRP3 have been proposed to be prognostic markers for the clinical outcomes of allogenic transplantation [92]. In addition to the strong genetic association between NLR polymorphisms/mutations and GVHD susceptibilities by human SNP analysis, animal experiments using NOD2-deficient mice suggested a role of NOD2 in GVHD pathophysiology. In contrast to TLRs, which drove donor T lymphocyte activation and severe GVHD outcome [32], the absence of NOD2 from the bone marrow transplant donor mice had no significant impact on the development of GVHD and did not regulate alloactivation of donor T cells. Instead, NOD2 deficiency in allo-bone marrow transplantation recipients caused increased GVHD occurrence in both MHC-mismatched and MHC-matched models [93, 94]. Furthermore, the proliferation and activation of donor T cells were enhanced by the absence of NOD2 from all bone marrow transplant recipients. This suggests that NOD2 plays a role in the regulation of host APCs, as evident by NOD2-deficient DCs displayed a higher activation status and increased ability to induce T cell proliferation during GVHD [93]. Several studies demonstrated a significant association between NOD2 SNPs and increased GVHD incidence/severity [6, 35, 88-91]. In contrast to these studies, several clinical findings did not find a significant impact of NOD2 SNPs on the incidence of GVHD [95-98]. Multiple factors could explain the conflicting results on the role of NOD2 and these may include differences in study population with NOD2 SNP frequency, overall incidence of GVHD, T cell activities, donor and recipient immune differences, and variation in environmental factors. Nonetheless, these data give new insight into the potential role of NOD2 in GVHD pathophysiology.
Many cytokines have been identified to be risk factors for predicting the outcome and severity of GVHD [99]. IL-1β is one of key inflammatory cytokines involved in GVHD pathophysiology and increased production of IL-1β is found to be associated with worse outcomes of GVHD [100]. Given that IL-1β is a major pro-inflammatory cytokine released following the activation of inflammasome, a genetic association study was conducted to evaluate whether mutations in NLRP genes are associated with GVHD pathogenecity. The genotypes of 133 patients undergoing HLA-identical sibling allogenic stem cell transplantation (allo-SCT) were analyzed and a strong association between common genetic variants in NLRP2 and NLRP3 genes and clinical outcome of HLA-identical allo-SCT was reported [92].
Despite increasing evidence, suggesting the importance of common variants in NOD2 or NLRP genes on the clinical outcome of GVHD, the mechanisms underlying these associations remain unidentified. Cellular damage or stress can ultimately lead to release of danger signals, which cause excessive immune-mediated tissue destruction, often found in acute GVHD. Adenosine-5-triphosphate (ATP) has been recognized as an endogenous danger signal during GVHD [4, 101, 102]. Extracellular ATP and its receptor P2X7R are expressed on several immune cell types and have been shown to be an important signal for activating NLRP3 inflammasome [78]. Increased concentration of ATP was reported during the development of GVHD and ATP-mediated stimulation of APCs led to increased expression of co-stimulatory molecules, such as CD80 and CD86 and further activated a variety of pro-inflammatory signaling pathways leading to expansion of donor T cells but reduction of regulatory T cells [101]. This data gives a new insight into the mechanisms by which danger signals enhance GVHD. Overall, studies characterizing the role of endogeneous or exogenous danger signals as well as their innate immune receptors in GVHD may hopefully help to develop a potential therapeutic target to bring a better clinical outcome for patients undergoing SOT or HCT.
Go to :
CONCLUSION
Innate immune receptors significantly contribute to the inflammatory processes that consequently lead to the recruitment of alloactivated T cells as well as tissue damage in GVHD target organs. Fig. 3 describes how the development of GVHD could be influenced by innate immune response. The generation of adaptive immunity can be triggered by activation of TLR-mediated APCs engulfing microbial pathogens or sensing danger signals in the peripheral tissues. Subsequently, APCs migrate to the draining lymph nodes to present the processed peptides to naïve T cells in the context of MHC molecules. Thus, TLRs provide signals necessary for this migration or could act directly on T cell function. Consequently, this could result in accelerating the development of severe GVHD.
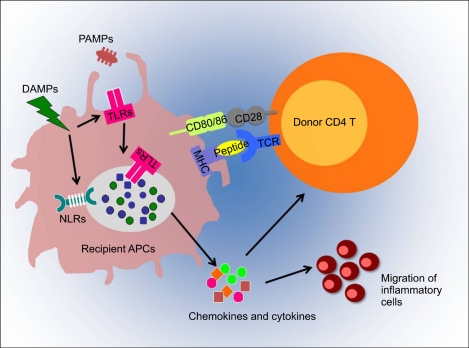 | Fig. 3Potential role of TLRs/NLRs in GVHD. GVHD occurs as a result of donor T lymphocyte activation reacting to antigens presented by recipient APCs. Recipient APCs, such as dendritic cells and macrophages, recognize PAMPs and DAMPs via TLRs/NLRs. Microbial pathogens or danger signals can be engulfed by phagocyotsis/endocytosis or through unidentified mechanisms, activate TLRs/NLRs on APCs. The ligation of TLRs leads to the maturation of APCs at the inflammatory site and enhances expression of co-stimulatory molecules and pro-inflammatory cytokines. This leads to a consequent migration of APCs to the draining lymph nodes. There, the APCs present alloantigens to naïve T cells, resulting in T cell activation and expansion. Activated T cells migrate back to the allograft and carry out their effector functions by attacking the graft. Thus, TLRs/NLRs-mediated antigen presentation may indirectly influence the alloreactivity of T cells or TLRs/NLRs may have a direct effect on T cells, thereby enhancing GVHD.
|
To date, most therapeutic and prophylactic efforts to prevent GVHD have been relied on immunosuppression mechanisms via T cell depletion or inhibition of T cell activation, proliferation, or effector function. Alternatively, immunosuppressive therapeutic strategies targeting key innate immune receptor signaling pathways may also decrease the severity and incidence of GVHD. Anti-IL-1β therapies have been successfully used in treating auto-inflammatory diseases [103] and the use of IL-1β antagonist to reduce the severity of GVHD has been investigated [104]. Further studies on the role of TLRs/NLRs control of subsequent adaptive immune response will enhance our knowledge of the overall host response to transplant grafts and hopefully lead to the development of strategies to modify this response and to improve clinical outcomes of SOT or HCT.
Go to :
 XML Download
XML Download