

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Fucoidan is a highly sulfated glycosaminoglycan, which has a molecular structure similar to that of heparin, and its antithrombotic effects in vitro have been widely reported [1-3]. Although fucoidan and heparin have similar structures, fucoidan exerts strong antiproliferative effects on smooth muscle cells (SMCs) [4] and also on heparin-resistant SMCs [5]. Furthermore, fucoidan can modulate the proliferation of fibroblasts [6].
The pathogenesis of atherothrombotic diseases such as stroke, myocardial infarction, and restenosis after angioplasty affects vascular SMC proliferation, human umbilical vascular endothelial cell (HUVEC) inflammation, and platelet aggregation [7, 8]. In addition, the major component of restenosis after stent implantation is intimal hyperplasia due to migration and proliferation of vascular SMCs from the media to the intima [9, 10].
In this study, we tried to investigate the effects and inhibitory mechanism of fucoidan on thrombosis, both in vitro and in vivo, as well as on vascular cells. Therefore, we first evaluated the antithrombotic effects of fucoidan on blood coagulation factors and platelet aggregation in vitro. We also examined the antithrombotic effect and mechanism of fucoidan in a mouse carotid artery thrombosis model because not many reports are available on the antithrombotic effects of fucoidan in vivo (i.e., in an artery injury model). Furthermore, we investigated the effects of fucoidan on SMCs and HUVECs, which play a key role in atherosclerotic vascular diseases. In particular, the change in the inflammatory factors in vascular cells was analyzed by Multiplex cytokine assay system.
Go to : 
MATERIALS AND METHODS
1. Ferric chloride (FeCl3)-induced mouse thrombosis model
Slc:ICR mice (6-8 weeks old males; weight, 30-33 g) were purchased from Jungang Animal Co., Seoul, Korea. Thrombosis was induced in mice using a previously described carotid artery injury model [11]. Three mice were used for each concentration. After an intraperitoneal injection containing both 2.5 mL/kg Zoletil (Virbac Animal Health Co., Carros, France) and 0.3 mL/kg Rompun (Bayer Korea, Anseong, Korea) for anesthesia, the skin on the upper central neck area was directly incised and the left common carotid artery was exposed. Carotid artery blood flow was measured with a miniature Doppler flow probe (diameter: 2 mm) for photoplethysmography (BioBud Inc., Seoul, Korea). Total occlusion was induced by applying a strip of filter paper (1×1 mm) saturated with 25% FeCl3 proximal to the surface of the carotid artery for 3 min. The carotid blood flow was monitored from time 0 (before the application of the FeCl3 paper) up to the time of occlusion. Various concentrations (0, 0.05, 0.07, and 0.1 mg/kg body weight) of fucoidan (Sigma Chemical Co., St. Louis, MO, USA) or high-molecular-weight heparin (100 KU, Grade I-A, Sigma Chemical Co) were intravenously injected just before blood flow monitoring. The concentrations of heparin were 0, 0.05, 0.07, 0.1, 0.13, 0.16, and 0.2 mg/kg body weight.
2. Platelet aggregation assay
The platelet aggregation assay was performed in human platelet-rich plasma (PRP). The platelet concentrate was diluted to 300,000 cells/µL of platelet-poor plasma (PPP). Ten microliters of samples (fucoidan, heparin, and phosphate-buffered saline [PBS]) was added to 450 µL of plasma and incubated for 2 min in an incubation well of the platelet aggregometer (Chrono-Log Co., Havertown, PA, USA) at 37℃. The final concentrations of fucoidan and heparin in this experiment were 0, 0.2, 0.4, 0.6, 0.8, and 1 µg/mL. The impedance was recorded, and ADP (20 µM) was added in order to initiate platelet aggregation. The inhibition of platelet aggregation was measured at the maximum aggregation response and the IC50 value was calculated by the least squares method.
3. Inhibition of thrombin and factor Xa by antithrombin in the presence of fucoidan or heparin
The anti-thrombin and anti-factor Xa activities of fucoidan were determined by chromogenic assay methods. In a 1.5 mL tube that contained thrombin (0.1 units/mL) or factor Xa (0.005 units/mL) in a buffer (pH 8.8) containing 50 mM Tris-HCl and 38 mM NaCl and antithrombin (0.1 µg/mL), various concentrations (0, 1, 5, 10, 50, 100, and 500 µg/mL) of fucoidan or heparin (180 IU/mg) were added, and the tubes were incubated at 37℃ for 20 min. After incubation, 25 µL of the solution from each 1.5 mL tube was transferred to a well of a 96-well plate. Then, 200 µL of 50 mM Tris-HCl (pH 8.8) buffer and 25 µL of thrombin substrate (0.3 mM/mL) or factor Xa substrate (0.3 mM/mL) were added to each well of the 96-well plate and incubated for 60 min at 37℃. After incubation, amidolytic activities were measured using a spectrophotometer at 405 nm.
4. Coagulation parameters
PPP was obtained by centrifugation of citrated human blood at 1,200×g for 15 min. The plasma was incubated with fucoidan dissolved in PBS (plasma : PBS=1 : 4), heparin dissolved in PBS, or only PBS for 2 min at 37℃. The plasma clotting times, activated partial thromboplastin time (aPTT) (Hyphen-BioMed, Andresy, France), prothrombin time (PT) (Hyphen-BioMed), and thrombin time (TT) (Hyphen-BioMed) were measured using the blood coagulometer Thrombotimer 2 (Behnk Elektronik, Norderstedt, Germany).
5. Rat aortic smooth muscle cell (RAoSMC) culture and proliferation assay
RAoSMCs were obtained from BioBud Inc. (Seoul, Korea). The cells were maintained in Dulbecco's modified Eagle's medium (DMEM, Gibco Life Technologies, Gaithersburg, MD, USA) containing 10% fetal bovine serum (Welgene Co., Daegu, Korea), 3.7% sodium bicarbonate, and 1% penicillin-streptomycin (10,000 IU/mL; Gibco Life Technologies). RAoSMCs were used between the fourth and eighth passages. The cells were incubated in 96-well plates (Corning Costar, Cambridge, MA, USA) at a density of 5×104 cells/well. After 24 h, the medium was changed with 100 µL fresh media containing several different concentrations (0, 50, 100, 500, and 1,000 µg/mL) of fucoidan and high-molecular-weight heparin. After changing the media, cells were further incubated for 24, 48, and 72 h at 37℃. Assays for cell proliferation were performed using a CCK-8 kit (Dojindo Molecular Technologies Inc, MA, USA). The samples were read at 450 nm using a spectrophotometer VERSAmax™ Tunable Microplate Reader (Molecular Devices Corp., CA, USA).
6. SMC migration assay
The effect of fucoidan on SMC migration was examined by performing a modified Boyden chamber assay in Transwell cell culture chambers using a polycarbonate membrane with 8 µm pores in a 6-well plate. RAoSMCs were suspended in serum-free DMEM at a concentration of 5×105 cells/mL. Cells were then pretreated with several concentrations of fucoidan and heparin for 30 min at 37℃. Serum-free DMEM was added to the lower compartment. Cell suspensions were added to the upper compartment and were then incubated for 6 h at 37℃. The cells were fixed and stained with crystal violet. The filter membranes were dried and the migrated cells were counted.
7. SMC adhesion assay
Ninety-six-well plates were precoated with vitronectin, fibronectin, and laminin (500 µg/mL) at 4℃ overnight. SMCs (3×105 cells/well) were preincubated with fucoidan and heparin for 30 min at 37℃. After incubation, the cells were transferred to each well and incubated for 1 h at 37℃ in 5% CO2. Unattached cells were removed by washing with PBS. Attached cells were fixed and stained with crystal violet. The plates were read using a spectrophotometer at 490 nm to determine the relative number of cells.
8. Mutiplex cytokine assays in HUVECs
HUVECs were plated onto 96-well cell culture plates coated with 0.5% gelatin. After 24 h, the serum-free media containing fucoidan and heparin were changed and the plate was incubated for 8 h at 37℃ in a CO2 atmosphere; the cells were then stimulated with tumor necrosis factor-alpha (TNFα) (10 ng/mL) for 16 h in an incubator. After incubation, supernatants were collected for the determination of the inflammatory cytokine levels using a Multiplex cytokine assay system (Luminex 100™ IS Total System, XMAP technology, Millipore, MA, USA). Sixteen kinds of inflammatory cytokines and/or chemokines (Eotaxin, granulocyte-macrophage colony-stimulating factor [GM-CSF], interferon (IFN)-α2, IFNγ, interleukin (IL)-10, IL-12, IL-1α, IL-1β, IL-1Rα, IL-2, IL-6, IP-10, monocyte chemoattractant protein (MCP)-1, RANTES, TNF-α, and TNF-β) were quantitatively determined.
Go to :
RESULTS
1. Inhibitory effect of fucoidan in a mouse carotid artery thrombosis model
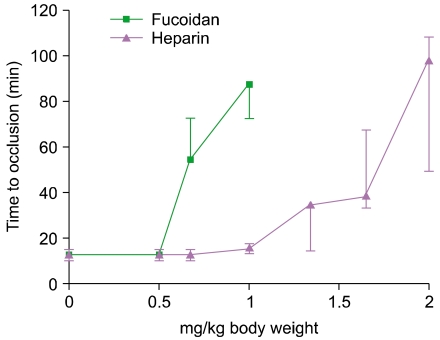
A FeCl3-induced mouse carotid artery occlusion model was developed and well optimized to evaluate the antithrombotic activity of anticoagulants and thrombolytic agents (Fig. 1). Before drug treatment, the average total occlusion time was determined as 12.5 min. Antithrombotic activity (effective dose, 50% [ED50]) was calculated as the concentration required to double the total occlusion time. The ED50 values of fucoidan and heparin were approximately 0.54 and 1.24 mg/kg body weight, respectively. Therefore, fucoidan showed approximately 2.3 times stronger antithrombotic effect than heparin in vivo (Fig. 2).
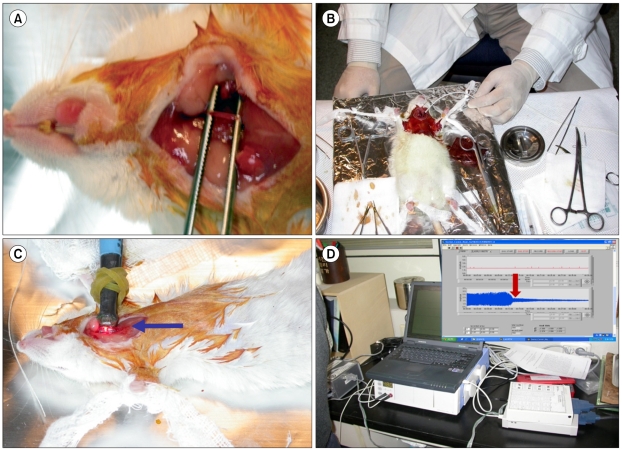 | Fig. 1Ferric chloride-induced mouse carotid artery thrombosis model. (A) Expose of left common carotid artery, (B) Induction of total occlusion by applying a filter paper saturated with 25% ferric chloride solution, (C) Monitoring of carotid blood flow with micro-optical probe, (D) Photoplethysmography of carotid blood flow until time to occlusion.
|
2. Inhibitory effect of fucoidan on platelet aggregation
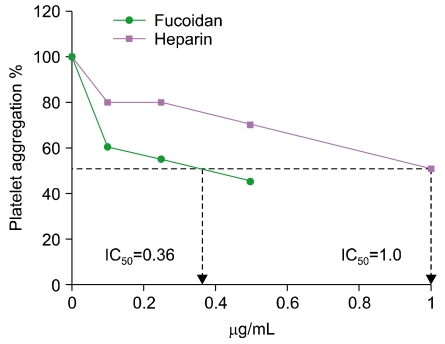
Fucoidan strongly inhibited ADP-induced human platelet aggregation in vitro in a concentration-dependent manner. The 50% inhibitory concentrations (IC50 values) of fucoidan and heparin for platelet aggregation were determined as 0.36 and 1.0 µg/mL, respectively (Fig. 3). Therefore, the inhibitory effect of fucoidan on platelet aggregation is approximately 2.8 times superior to that of heparin.
3. Anti-thrombin and anti-factor Xa activities of fucoidan
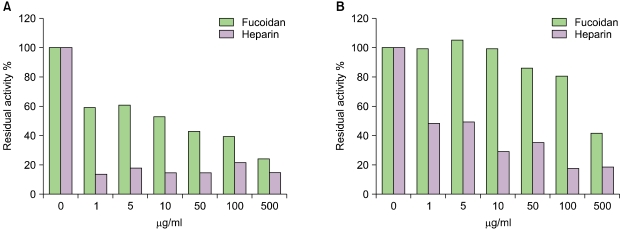
The anti-thrombin and anti-factor Xa activities of fucoidan were interestingly weaker than those of heparin (Fig. 4), although the overall antithrombotic effect of fucoidan in vivo was even more potent than that of heparin (data not shown).
4. Effects of fucoidan on PT, aPTT, and TT
The anticoagulation activities of fucoidan, as determined by aPTT (Table 1), PT (data not shown), and TT (data not shown), were revealed to be dose dependent, although heparin showed considerably higher inhibitory activity on PT, aPTT, and TT than fucoidan.

5. Inhibitory effects of fucoidan on SMC proliferation, migration, and adhesion
We tested the inhibitory effects of fucoidan on RAoSMC proliferation, migration, and adhesion. In proliferation assays, we found that fucoidan had a stronger inhibitory effect on SMC proliferation than heparin (Fig. 5). In addition, fucoidan inhibited SMC migration and SMC adhesion to extracellular matrix to a greater extent than heparin (data not shown). Specially, fucoidan inhibited SMC adhesion to laminin in a dose-dependent manner.
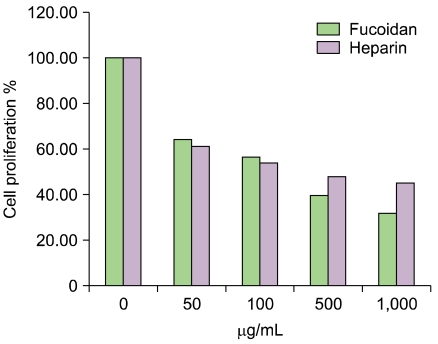 | Fig. 5The inhibitory effect of fucoidan on SMC proliferation. The antiproliferative effect of fucoidan on SMC was stronger than that of heparin. Cell proliferation was inhibited at 50 µg/mL of both fucoidan and heparin. Above a concentration of 500 µg/mL, fucoidan showed a stronger antiproliferative effect on SMCs than heparin. *P<0.01
|
6. Effects of fucoidan on cytokine production in HUVECs
Sixteen kinds of cytokines were simultaneously determined by the Multiplex assay system. We confirmed that the levels of some specific inflammatory cytokines and chemokines were reduced in HUVECs when we treated the cells with fucoidan in the presence of TNF-α. Specifically, fucoidan treatment reduced the amounts of GM-CSF, IL-6, MCP-1, and RANTES (Fig. 6). In addition, fucoidan showed a stronger inhibitory effect on the proinflammatory cytokines than heparin.
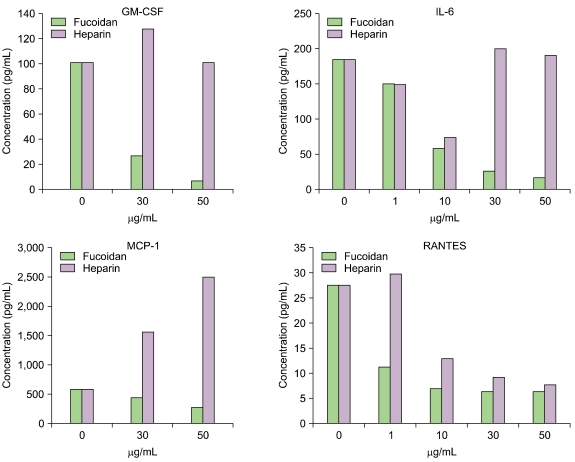
Go to :
DISCUSSION
The purpose of this study was to investigate the effects and mechanism of fucoidan on thrombosis both in vitro and in vivo and in vascular cells such as SMCs and endothelial cells. Because many reports are not available on the antithrombotic effect of fucoidan in vivo, we optimized a FeCl3-induced mouse carotid artery occlusion model by the modification of a previous model [11] to evaluate the antithrombotic effects of fucoidan.
The antithrombotic activities of fucoidan in vitro were similar to those reported by other groups [12-17]. However, in in vivo studies using a mouse thrombosis model, fucoidan showed stronger antithrombotic activities than heparin. To define the antithrombotic mechanism of fucoidan, we examined the effects of fucoidan on blood coagulation factors and platelet aggregation in vitro. The anti-thrombin and anti-factor Xa activities of fucoidan in vitro were weaker than those of heparin (Fig. 4), suggesting that the stronger antithrombotic activities of fucoidan in vivo might be caused by its binding with heparin cofactor II rather than with antithrombin. The potent anti-thrombin and anti-factor Xa activities of heparin in vitro may be due to the increase in its binding affinity with antithrombin.
The effects of fucoidan and heparin on coagulation time have also been examined by aPTT, PT, and TT assays in human PPP. The anticoagulant activity of heparin was much stronger than fucoidan in this experiment. Therefore, it could be concluded that the antithrombotic mechanism of fucoidan is different from that of heparin.
One of the major causes of restenosis after stenting is SMC proliferation and migration [9, 10]. A previous report demonstrated that the mechanism of the antimitogenic effect of fucoidan was different from that of heparin, which also inhibits SMC growth [18]. As expected, fucoidan inhibited proliferation and migration of SMCs to a greater extent than heparin [19].
It has recently been accepted that atherosclerosis and restenosis are an inflammatory process [20, 21]. To examine the effects of fucoidan and heparin on the production of cytokines in HUVECs, we determined the change in the inflammatory cytokines in HUVECs treated with fucoidan by the Multiplex assay system. Interestingly, the levels of several proinflammatory cytokines such as GM-CSF, IL-6, MCP-1, and RANTES were dramatically reduced in HUVECs treated with fucoidan.
In conclusion, fucoidan showed a stronger antithrombotic effect than heparin in vivo, antiproliferative effect on SMCs, and anti-inflammatory effect on HUVECs. Considering these results together, we can suggest that fucoidan may be useful for the prevention of cardiovascular diseases.
Go to :
 XML Download
XML Download