

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Introduction
This study examined the regulatory mechanism underlying the meal-induced changes in the hypothalamic-pituitary-adrenal gland (HPA) axis activity.
Materials and Methods
Male Sprague-Dawley rats (250–300 g) were hired for two different experiments as follows; 1) rats received either 8% sucrose or 0.2% saccharin ad libitum after 48 h of food deprivation with the gastric fistula closed (real feeding) or opened (sham feeding). 2). rats received 5 ml of intra-oral infusion with 0.2% saccharin or distilled water after 48 h of food deprivation. One hour after food access, all rats were sacrificed by a transcardiac perfusion with 4% paraformaldehyde. The brains were processed for c-Fos immunohistochemistry and the cardiac blood was collected for the plasma corticosterone assay.
Results
Real feedings with sucrose or saccharin and sham feeding saccharin but not sucrose, following food deprivation decreased the plasma corticosterone level. c-Fos expression in the nucleus tractus of solitarius (NTS) of the fasted rats was increased by the consumption of sucrose but not saccharin, regardless of the feeding method. On the other hand, the consumption of sucrose or saccharin with real feeding but not the sham, induced c-Fos expression in the paraventricular nucleus (PVN) of the fasted rats. The intra-oral infusion with saccharin or water decreased the plasma corticosterone level of the fasted rats. Intra-oral water infusion increased c-Fos expression in both the PVN and NTS, but saccharin only in the NTS in the fasted rats.
Conclusion
Neither restoration of the fasting-induced elevation of plasma corticosterone nor the activation of neurons in the PVN and NTS after refeeding requires the palatability of food or the post-ingestive satiety and caloric load. In addition, neuronal activation in the hypothalamic PVN may not be an implication in the restoration of the fasting-induced elevation of the plasma corticosterone by oropharyngeal stimuli of palatable food.
References
1. Isse T, Ueta Y, Serino R, Noguchi J, Yamamoto Y, Nomura M, et al. Effects of leptin on fasting-induced inhibition of neuronal nitric oxide synthase mRNA in the paraventricular and supraoptic nuclei of rats. Brain Res. 1999; 846:229–35.

2. Nelson RJ, Kriegsfeld LJ, Dawson VL, Dawson TM. Effect of nitric oxide on neuroendocrine function and behavior. Front Neuroendocrinol. 1997; 18:463–91.
3. Watts AG. Understanding the neural control of ingestive behaviors: helping to separate cause from effect with dehydration-associated anorexia. Horm Behav. 2000; 37:261–83.
4. Antoni FA. Hypothalamic control of adrenocorticotropin secretion: advances since the discovery of 41-residue corticotropin-releasing factor. Endocr Rev. 1986; 7:351–78.
5. Whitnall MH. Regulation of the hypothalamic corticotropin-releasing hormone neurosecretory system. Prog Neurobiol. 1993; 40:573–629.
6. Kiss A, Jezova D, Aguilera G. Activity of the hypothalamic pituitary adrenal axis and sympathoadrenal system during food and water deprivation in the rat. Brain Res. 1994; 663:84–92.
7. van Haasteren GA, Linkels E, van Toor H, Klootwijk W, Kaptein E, de Jong FH, et al. Effects of longterm food reduction on the hypothalamus-pituitary-thyroid axis in male and female rats. J Endocrinol. 1996; 150:169–78.
8. Yoshihara T, Honma S, Katsuno Y, Honma K. Dissociation of paraventricular NPY release and plasma corticosterone levels in rats under food deprivation. Am J Physiol. 1996; 271:E239–45.
9. Kim YM, Lee JY, Choi SH, Kim DG, Jahng JW. RU486 blocks fasting-induced decrease of neuronal nitric oxide synthase in the rat paraventricular nucleus. Brain Res. 2004; 1018:221–6.
10. Timofeeva E, Picard F, Duclos M, Deshaies Y, Richard D. Neuronal activation and corticotropin-releasing hormone expression in the brain of obese (fa/fa) and lean (fa/?) Zucker rats in response to refeeding. Eur J Neurosci. 2002; 15:1013–29.
11. Jahng JW, Lee JY, Yoo SB, Kim YM, Ryu V, Kang DW, et al. Refeeding-induced expression of neuronal nitric oxide synthase in the rat paraventricular nucleus. Brain Res. 2005; 1048:185–92.
12. Fraser KA, Davison JS. Meal-induced c-fos expression in brain stem is not dependent on cholecystokinin release. Am J Physiol. 1993; 265:R235–9.
13. Rinaman L, Baker EA, Hoffman GE, Stricker EM, Verbalis JG. Medullary c-fos activation in rats after ingestion of a satiating meal. Am J Physiol. 1998; 275:R262–68.
14. Fraser KA, Raizada E, Davison JS. Oral-pharyngeal-esophageal and gastric cues contribute to meal-induced c-fos expression. Am J Physiol. 1995; 268:R223–30.
15. Phillips RJ, Powley TL. Gastric volume rather than nutrient content inhibits food intake. Am J Physiol. 1996; 271:R766–9.
16. McCann MJ, Rogers RC. Impact of antral mechanoreceptor activation on the vago-vagal reflex in the rat: functional zonation of responses. J Physiol. 1992; 453:401–11.
17. Raybould HE, Gayton RJ, Dockray GJ. CNS effects of circulating CCK8: involvement of brainstem neurones responding to gastric distension. Brain Res. 1985; 342:187–90.
18. Harrer MI, Travers SP. Topographic organization of Fos-like immunoreactivity in the rostral nucleus of the solitary tract evoked by gustatory stimulation with sucrose and quinine. Brain Res. 1996; 711:125–37.
19. Norgren R. Gustatory system. Paxinos G, editor. The Rat Nervous System. 2nd ed.San Diego: Academic Press;1995. p. 751–71.
20. Yamamoto T. Electrophysiology of CTA. Bures J, Bermu ′ dez-Rattoni F, Yamamoto T, editors. Conditioned taste aversion: memory of a special kind. 1st ed.New York: Oxford Univ. Press;1998. p. 76–107.
21. Pritchard TC, Hamilton RB, Norgren R. Projections of the parabrachial nucleus in the old world monkey. Exp Neurol. 2000; 165:101–17.
22. Yamamoto T, Matsuo R, Kiyomitsu Y, Kitamura R. Taste responses of cortical neurons in freely ingesting rats. J Neurophysiol. 1989; 61:1244–58.
23. Kobayakawa T, Ogawa H, Kaneda H, Ayabe-Kanamura S, Endo H, Saito S. Spatiotemporal analysis of cortical activity evoked by gustatory stimulation in humans. Chem Senses. 1999; 24:201–9.
24. Bermu ′ dez-Rattoni F. Molecular mechanisms of taste-recognition memory. Nat Rev Neurosci. 2004; 5:209–17.
25. Tsukamoto G, Adachi A. Neural responses of rat area postrema to stimuli producing nausea. J Auton Nerv Syst. 1994; 49:55–60.
26. Hermann GE, Kohlerman NJ, Rogers RC. Hepatic-vagal and gustatory afferent interactions in the brainstem of the rat. J Auton Nerv Syst. 1983; 9:477–95.
27. Cechetto DF, Saper CB. Evidence for a viscerotopic sensory representation in the cortex and thalamus in the rat. J Comp Neurol. 1987; 262:27–45.
28. Yoo SB, Lee JH, Ryu V, Jahng JW. Ingestion of non-caloric liquid diet is sufficient to restore plasma corticosterone level, but not to induce the hypothalamic c-Fos expression in food-deprived rats. Nutr Neurosci. 2007; 10:261–7.
29. Weingarten HP, Watson SD. Sham feeding as a procedure for assessing the influence of diet palatability on food intake. Physiol Behav. 1982; 28:401–7.
30. Conover KL, Shizgal P. Competition and summation between rewarding effects of sucrose and lateral hypothalamic stimulation in the rat. Behav Neurosci. 1994; 108:537–48.
31. Davis JD, Smith JP, Kung TM. Abdominal vagotomy alters the structure of the ingestive behavior of rats ingesting liquid diets. Behav Neurosci. 1994; 108:767–79.
32. Nissenbaum JW, Sclafani A. Sham feeding response of rats to polycose and sucrose. Neurosci Biobehav Rev. 1987; 11:215–22.
33. Emond MH, Weingarten HP. Fos-like immunoreactivity in vagal and hypoglossal nuclei in different feeding states: a quantitative study. Physiol Behav. 1995; 58:459–65.
34. Paxinos G, Watson C. The rat brain in Stereotaxic Coordinates. 2nd ed.San Diego: Academic Press;1986.
35. Ulrich-Lai YM, Ostrander MM, Thomas IM, Packard BA, Furay AR, Dolgas CM, et al. Daily limited access to sweetened drink attenuates hypothalamic-pituitary-adrenocortical axis stress responses. Endocrinology. 2007; 148:1823–34.
36. Suchecki D, Antunes J, Tufik S. Palatable solutions during paradoxical sleep deprivation: reduction of hypothalamic-pituitary-adrenal axis activity and lack of effect on energy imbalance. J Neuroendocrinol. 2003; 15:815–21.
37. Sclafani A, Nissenbaum JW. On the role of the mouth and gut in the control of saccharin and sugar intake: a reexamination of the sham-feeding preparation. Brain Res Bull. 1985; 14:569–76.
38. Sclafani A, Nissenbaum JW. Is gastric sham feeding really sham feeding? Am J Physiol. 1985; 248:R387–90.
39. Grill HJ, Norgren R. The taste reactivity test. I. Mimetic responses to gustatory stimuli in neurologically normal rats. Brain Res. 1978; 143:263–79.
40. Berridge KC. Modulation of taste affect by hunger, caloric satiety, and sensory-specific satiety in the rat. Appetite. 1991; 16:103–20.
41. Wacker J, Dillon DG, Pizzagalli DA. The role of the nucleus accumbens and rostral anterior cingulate cortex in anhedonia: integration of resting EEG, fMRI, and volumetric techniques. Neuroimage. 2009; 46:327–37.
42. Pizzagalli DA, Holmes AJ, Dillon DG, Goetz EL, Birk JL, Bogdan R, et al. Reduced caudate and nucleus accumbens response to rewards in unmedicated individuals with major depressive disorder. Am J Psychiatry. 2009; 166:702–10.
43. Shirayama Y, Chaki S. Neurochemistry of the nucleus accumbens and its relevance to depression and antidepressant action in rodents. Curr Neuropharmacol. 2006; 4:277–91.
44. Bruchas MR, Xu M, Chavkin C. Repeated swim stress induces kappa opioid-mediated activation of extracellular signal-regulated kinase 1/2. Neuroreport. 2008; 19:1417–22.
45. Kalivas PW, Duffy P. Selective activation of dopamine transmission in the shell of the nucleus accumbens by stress. Brain Res. 1995; 675:325–8.
46. Campioni MR, Xu M, McGehee DS. Stress-induced changes in nucleus accumbens glutamate synaptic plasticity. J Neurophysiol. 2009; 101:3192–8.
47. Lucas LR, Wang CJ, McCall TJ, McEwen BS. Effects of immobilization stress on neurochemical markers in the motivational system of the male rat. Brain Res. 2007; 1155:108–15.
48. Naimi N, Rivest S, Racotta I, Richard D. Neuronal activation of the hypothalamic magnocellular system in response to oropharyngeal stimuli in the rat. J Neuroendocrinol. 1997; 9:329–40.
49. Berthoud HR. Multiple neural systems controlling food intake and body weight. Neurosci Biobehav Rev. 2002; 26:393–428.
50. Yamamoto T, Sawa K. Comparison of c-fos-like immunoreactivity in the brainstem following intraoral and intragastric infusions of chemical solutions in the rats. Brain Res. 2000; 866:144–51.
Fig. 1.
Plasma corticosterone levels. Rats received ad libitum access to either 8% sucrose or 0.2% sodium saccharin for 1 h with gastric fistula closed (real feeding) or opened (sham feeding) following 48 h of food deprivation. Cardiac blood was collected immediately after exposing the heart for perfusion, and the plasma level of corticosterone was analyzed by radioimmunoassay.(FD: 48 h food deprivation) *P<0.05 vs. FD, Data is presented by means±S.E.
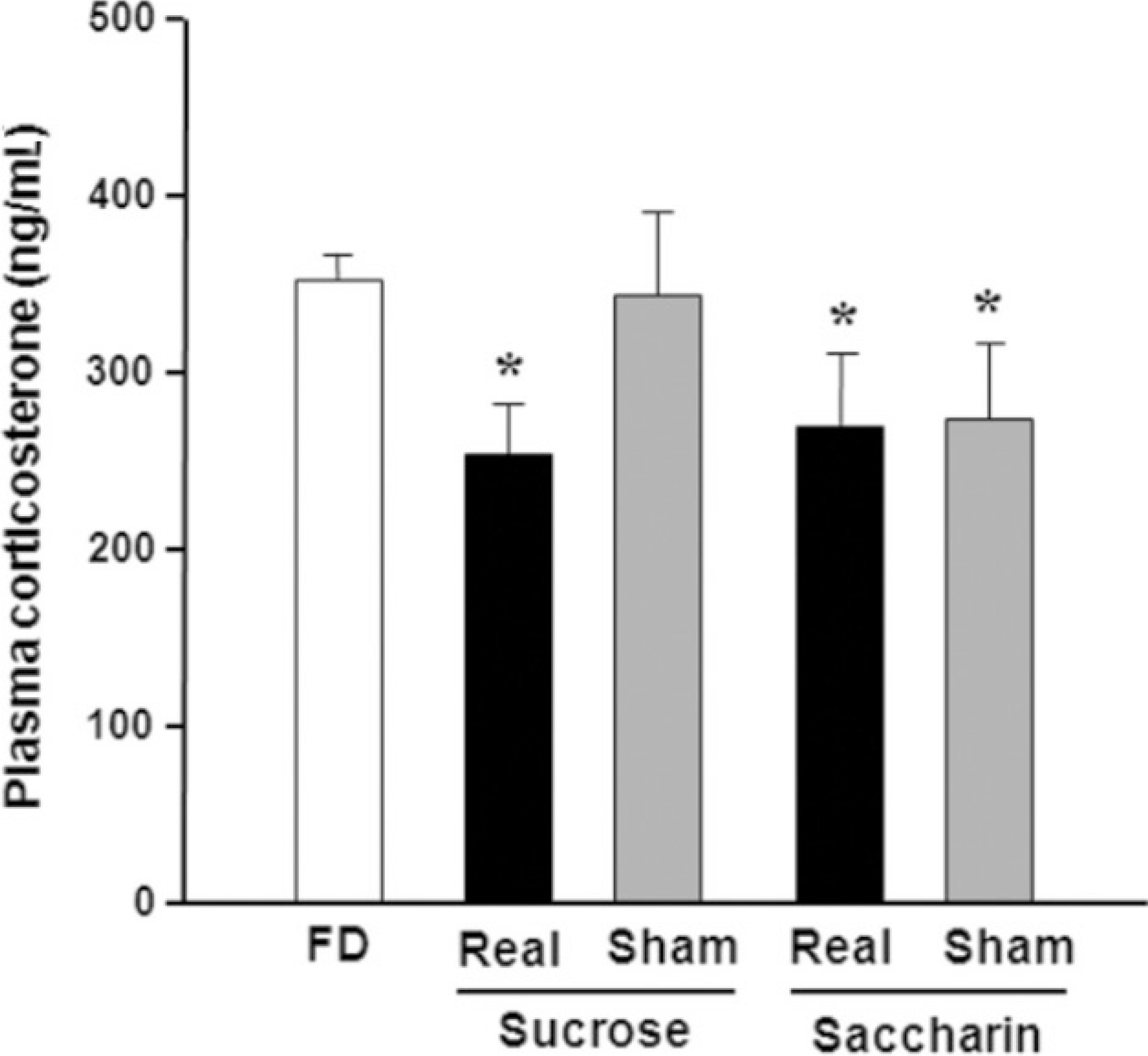
Fig. 2.
c-Fos immunohistochemistry in the PVN and the NTS. Rats were transcardiac perfused with 4 % PFA immediately after 1 h of ad libitum access to either 8% sucrose or 0.2% sodium saccharin for 1 h with gastric fistula closed (real feeding) or opened (sham feeding) following 48 h of food deprivation. Distinctive black dots represent c-Fos immunoreactivity (−ir) nuclei. A-E. PVN, f-j. caudal NTS (c-NTS), K-O. intermediate NTS (i-NTS). (FD: 48 h food deprivation, R/Suc: 1 h real refeeding with sucrose, S/Suc: 1 h sham refeeding with sucrose, R/Sac: 1 h real refeeding with saccharin, S/Sac: 1 h sham refeeding with saccharin, AP: area postrema, 3v: 3rd ventricle, 4v: 4th ventricle) Scale bars: 200μm.
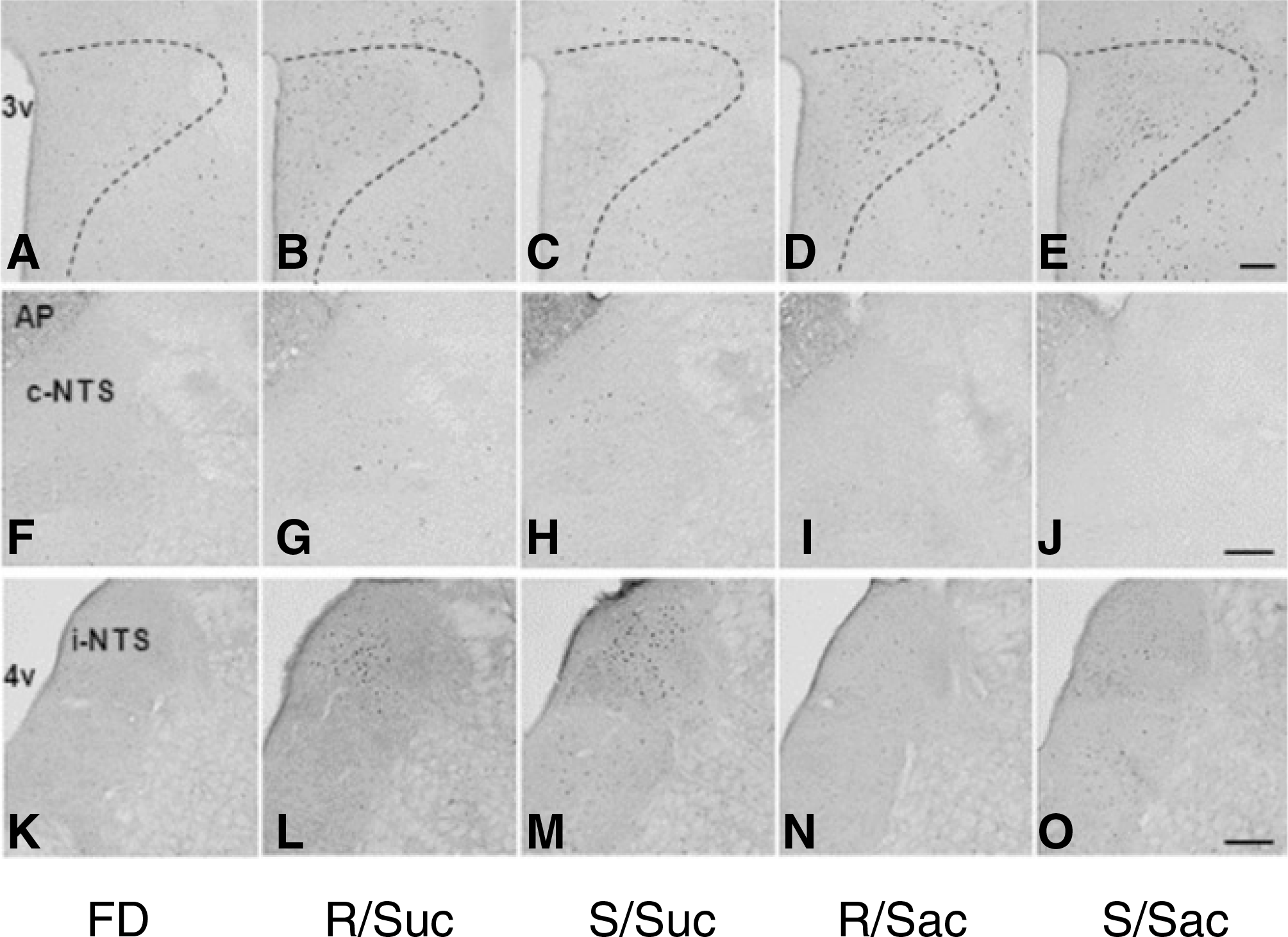
Fig. 3.
The number of c-Fos-immunoreactive (−ir) nuclei in the PVN (A) and NTS (B).(FD: 48 h food deprivation) *P<0.05, **P<0.01, ***P<0.001 vs. FD, Data is presented by means±S.E.
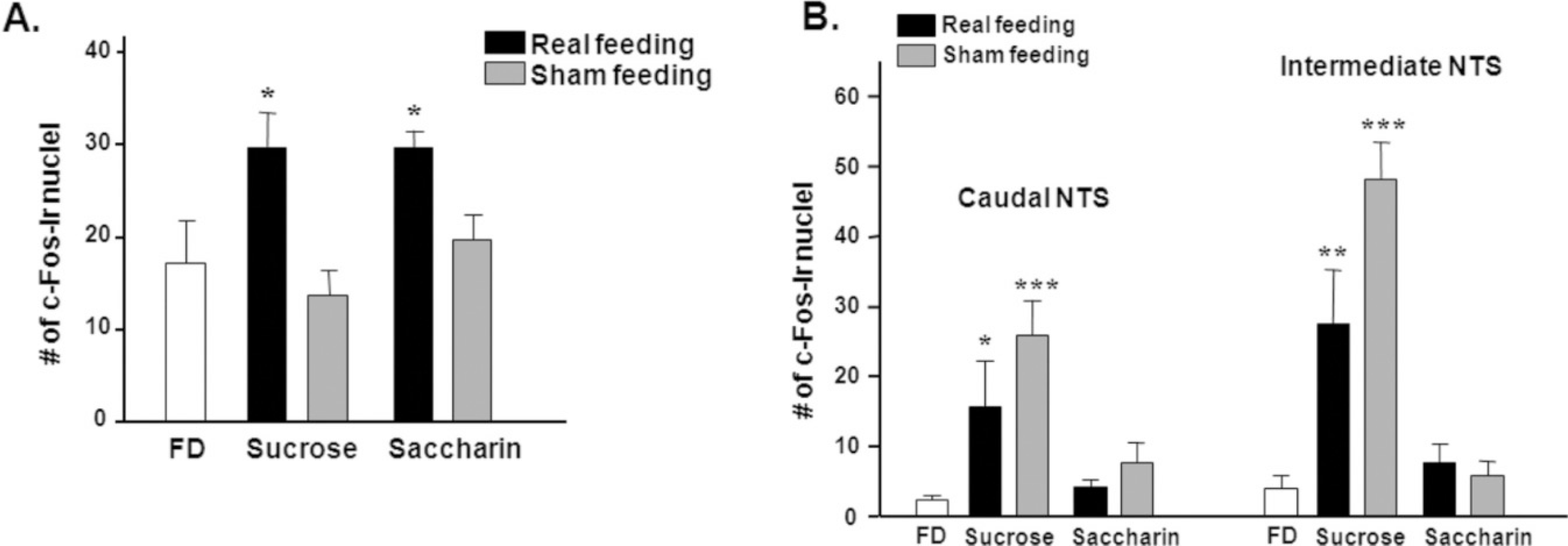
Fig. 4.
Plasma corticosterone levels. Rats received intra-oral infusion with saccharin or distilled water for 10 min following 48 h of food deprivation, and then were sacrificed for analyzing plasma level of corticosterone.(FD: 48 h food deprivation, Sac: intra-oral infusion with 0.2% saccharin, DW: intra-oral infusion with distilled water) *P<0.05, **P<0.01 vs. FD, Data is presented by means±S.E.
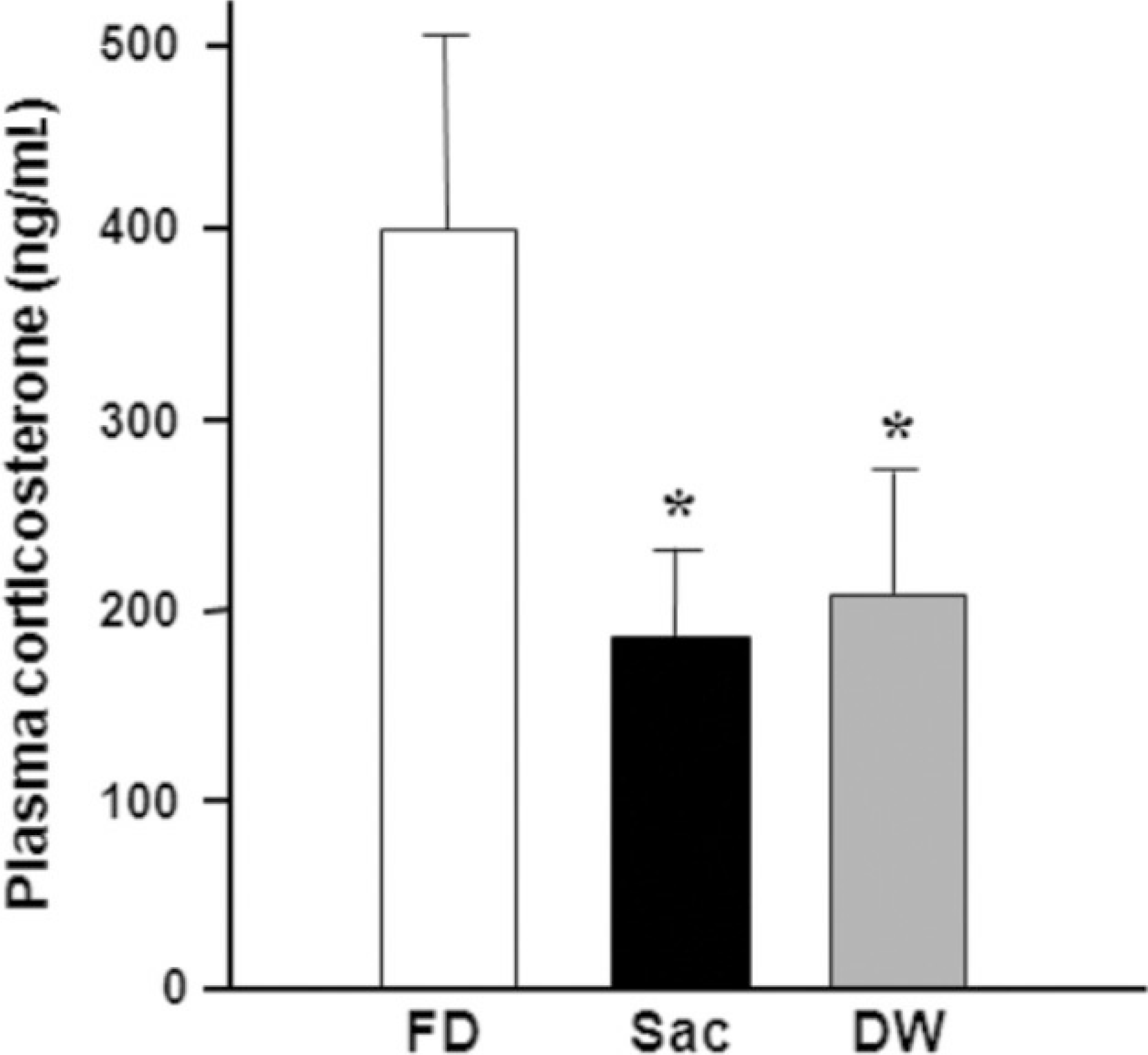
Fig. 5.
The number of c-Fos-immunoreactive (−ir) nuclei in the PVN (A) and NTS (B).(FD: 48 h food deprivation, Sac: intraoral infusion with 0.2% saccharin, DW: intra-oral infusion with distilled water) *P<0.05, **P<0.01 vs. FD, Data is presented by means±S.E.
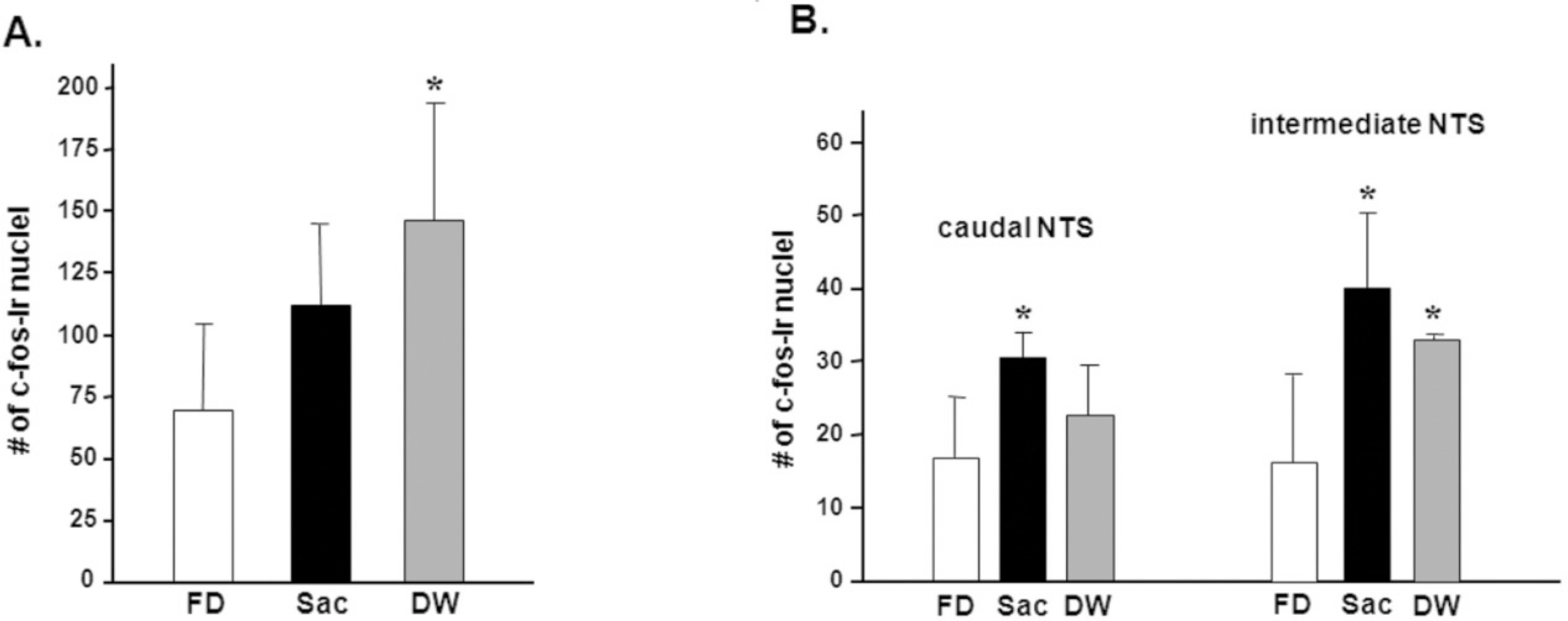
Table 1.
Liquid intake during 1 h of refeeding period following 48 h of food deprivation
| Groups | R/Suc | S/Suc | R/Sac | S/Sac |
|---|---|---|---|---|
| Liquid Intake (g) | 15.236±1.193 | 12.964±1.690 | 11.566±2.191* | 6.393±1.271 |
 XML Download
XML Download