

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Abstract
Introduction
Mammalian tooth pulp is densely innervated by sensory nerves that are mostly C fibers and A delta fibers. However, there is evidence suggesting that many unmyelinated axons in the pulp are in fact parent meylinated axons. Immunohistochemical staining for neurofilament protein 200 kDa (NFP200) was performed to identify the demyelinated but parent myelinated axons.
Materials and Methods
The pulp was removed from healthy premolars and 3rd molars extracted from juveniles and adults undergoing orthodontic treatment, and immunohistochemical staining were applied with NPF200 antibodies, which specifically dye myelinated axons. The specimens underwent an electron microscopy examination with diaminobenzidine (DAB) immunostaining after observation and analysis by fluorescence and confocal laser scanning microscopy.
Results
The NPF200 immuno-positive axons in the radicular pulp areas were observed as bundles of many nerve fibers. Many small bundles were formed with fewer axons when firing to the coronal pulp areas and then reachrd a different direction. In the radicular pulp, unmyelinated axons and myelinated axons were present together. However, in the coronal pulp, unmyelinated axons were most common and NFP200 immuno-positive unmyelinated axons with a larger diameter than those in the radicular pulp were observed more frequently. On the other hand, most of the immuno-positive unmyelinated fibers were similar in size to that of typically well-known unmyelinated fibers.
Go to : 
References
1. Byers MR. Detal sensory receptors. Int Rev Neurobiol. 1984; 25:39–94.
2. Hildebrand C, Fried K, Tuisku F, Johansson CS. Teeth and tooth nerves. Prog Neurobiol. 1995; 45:165–222.

3. Holland GR, Robinson PP. The number and size of axons at the apex of the cat's canine tooth. Anat Rec. 1983; 205:215–22.
4. Reader A, Foreman DW. An ultrastructural quantitative investigation of human intradental innervation. J Endod. 1981; 7:493–9.
5. Holje L, Hildebrand C, Fried K. Proportion of unmyelinated axons in the rat inferior alveolar nerve and mandibular molar pulps after neonatal administration of capsaicin. Brain Res. 1983; 266:133–6.
6. Horiuchi H. A study of pulp-nerve excitation through a silver-wire electrode. J Dent Res. 1965; 44:1257–63.
7. Lisney SJ. Some anatomical and electrophysiological properties of tooth-pulp afferents in the cat. J Physiol. 1978; 284:19–36.
8. Cadden SW, Lisney SJ, Matthews B. Thresholds to electrical stimulation of nerves in cat canine tooth-pulp with A beta-, A delta- and C-fibre conduction velocities. Brain Res. 1983; 261:31–41.
9. Fried K, Hildebrand C. Axon number and size distribution in the developing feline inferior alveolar nerve. J Neurol Sci. 1982; 53:169–80.
10. Johansson CS, Hildebrand C, Povlsen B. Anatomy and developmental chronology of the rat inferior alveolar nerve. Anat Rec. 1992; 234:144–52.
11. Paik SK, Park KP, Lee SK, Ma SK, Cho YS, Kim YK, et al. Light and electron microscopic analysis of the somata and parent axons innervating the rat upper molar and lower incisor pulp. Neuroscience. 2009; 162:1279–86.
12. Holland GR. Fibre numbers and sizes in the inferior alveolar nerve of the cat. J Anat. 1978; 127:343–52.
13. Hirvonen TJ. A quantitative electron-microscopic analysis of the axons at the apex of the canine tooth pulp in the dog. Acta Anat (Basel). 1987; 128:134–9.
14. Nair PN, Luder HU, Schroeder HE. Number and size-spectra of myelinated nerve fibers of human premolars. Anat Embryol (Berl). 1992; 186:563–71.
15. Naftel JP, Bernanke JM, Qian XB. Quantitative study of the apical nerve fibers of adult and juvenile rat molars. Anat Rec. 1994; 238:507–16.
16. Elder GA, Friedrich VL Jr, Bosco P, Kang C, Gourov A, Tu PH, et al. Absence of the mid-sized neurofilament subunit decreases axonal calibers, levels of light neurofilament (NF-L), and neurofilament content. J Cell Biol. 1998; 141:727–39.
17. Lawson SN, Waddell PJ. Soma neurofilament immunoreactivity is related to cell size and fibre conduction velocity in rat primary sensory neurons. J Physiol. 1991; 435:41–63.
18. Dirajlal S, Pauers LE, Stucky CL. Differential response properties of IB(4)-positive and -negative unmyelinated sensory neurons to protons and capsaicin. J Neurophysiol. 2003; 89:513–24.
19. Amaya F, Decosterd I, Samad TA, Plumpton C, Tate S, Mannion RJ, et al. Diversity of expression of the sensory neuron-specific TTX-resistant voltage-gated sodium ion channels SNS and SNS2. Mol Cell Neurosci. 2000; 15:331–42.
20. Beland B, Fitzgerald M. Mu- and delta-opioid receptors are downregulated in the largest diameter primary sensory neurons during postnatal development in rats. Pain. 2001; 90:143–50.
21. Bennett DL, Michael GJ, Ramachandran N, Muson JB, Averill S, Yan Q, et al. A distinct subgroup of small DRG cells express GDNF receptor components and GDNF is protective for these neurons after nerve injury. J Neurosci. 1998; 18:3059–72.
22. Benn SC, Costigan M, Tate S, Fitzgerald M, Woolf CJ. Developmental expression of the TTX-resistant voltage-gated sodium channels Nav1.8 (SNS) and Nav1.9 (SNS2) in primary sensory neurons. J Neurosci. 2001; 21:6077–85.
23. Naftel JP, Richards LP, Pan M, Bernanke JM. Course and composition of the nerves that supply the mandibular teeth of the rat. Anat Rec. 1999; 256:433–47.
Go to :
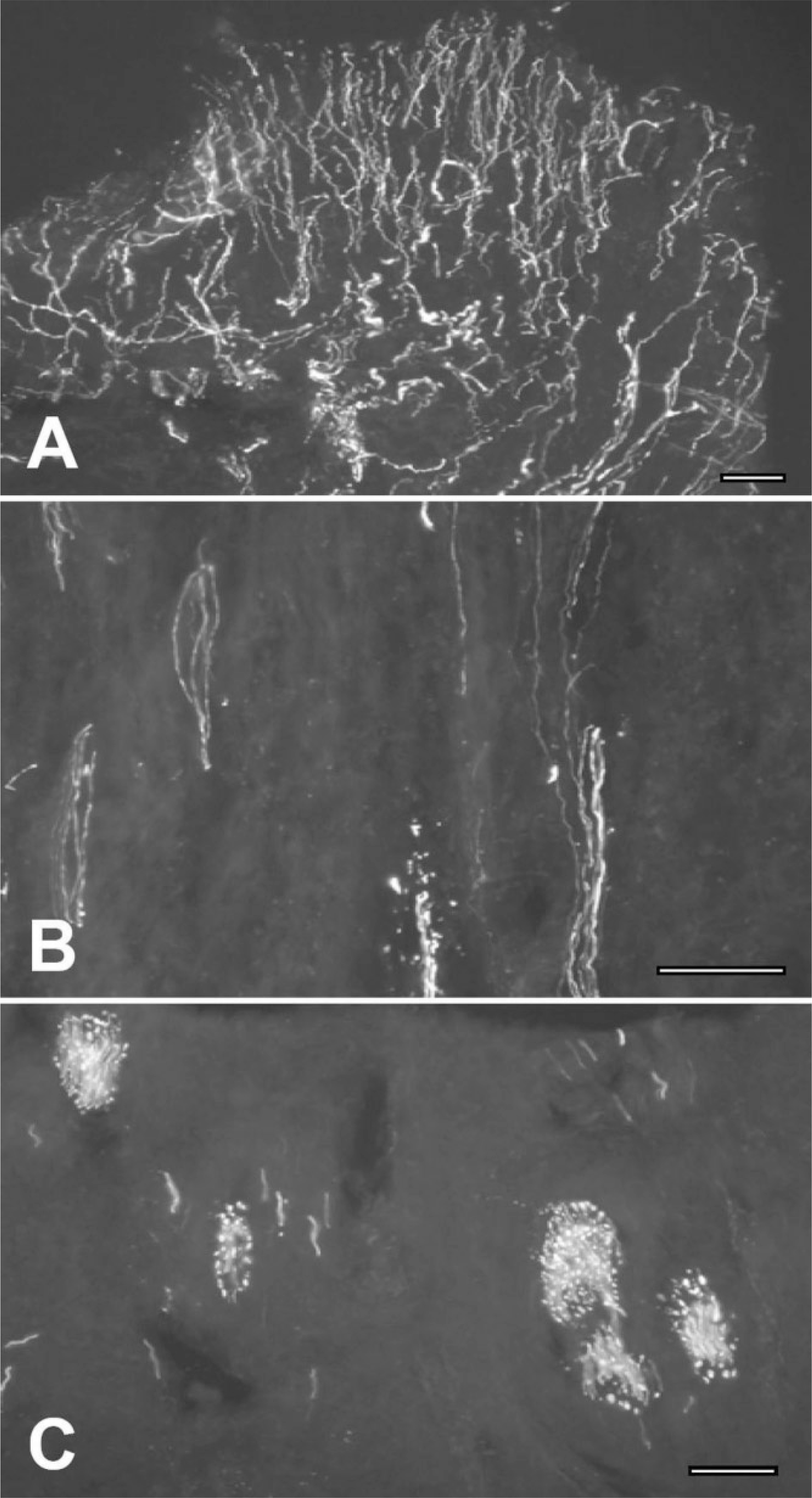 | Fig. 1.Immunofluorescence staining on human molar pulp showing the distribution of neurofilament protein (NFP) 200-immunopositive (+) nerves. A. NFP200+ nerves are densely appeared in subodontoblastic plexus (SOP) of Raschow of coronal pulp. B, C. NFP200+ nerve bundles C in root portion branch off and project to their termination sites. The scale bar is 50 μm in A-C. |
 | Fig. 2.Immunoperoxidase staining for electron microscopic observations on the neurofilament protein (NFP) 200-immunopositive nerves of human molar pulp. A, B. Many NFP200-immunopositive (+) nerves terminate in subodontoblastic plexus (SOP) of coronal pulp. Some nerves are also passing through odontoblast (OB) layer. C, D. Nerve bundles travel the root portion C and make branches before projecting to SOP of Raschow or OB layer. The scale bar is 100 μm in A and 50 μm in B-D. |
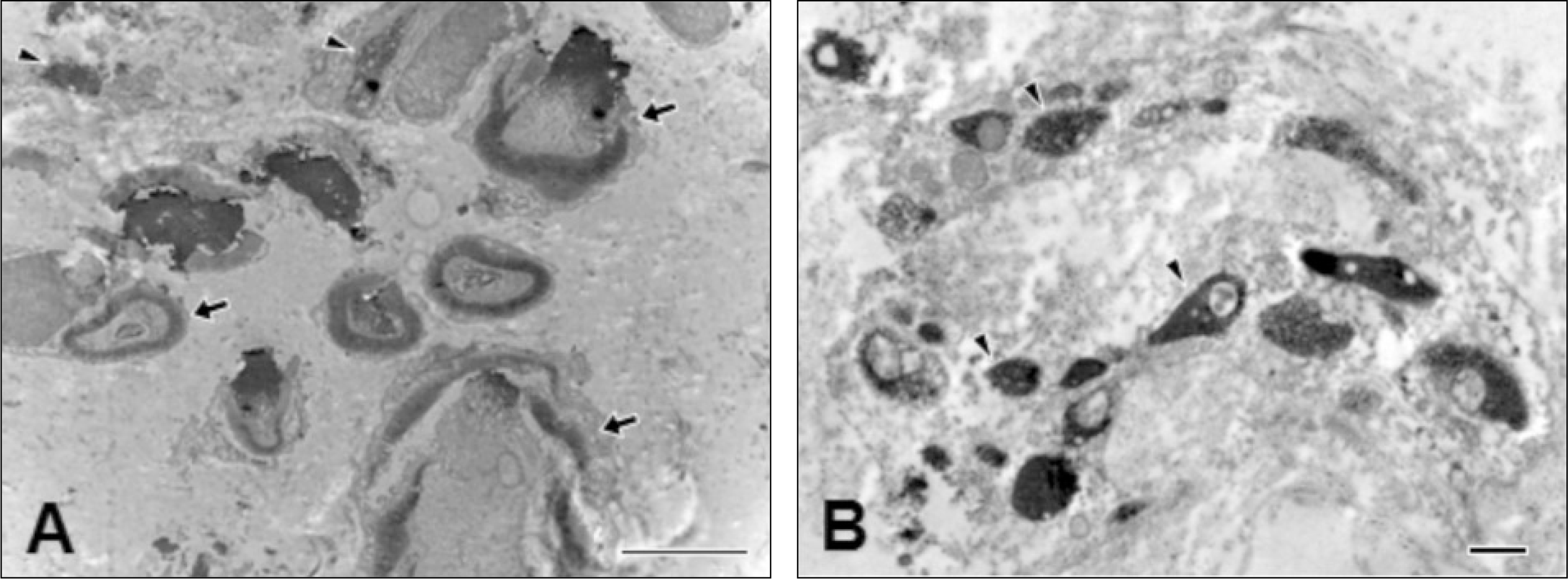 | Fig. 3.Electron microscopic observations on the neurofilament protein (NFP) 200-immunopositive (+) nerves in root and crown portions. A. NFP200+ myelinated (arrows) and unmyelinated (arrowheads) nerves are frequently observed in root portion. B. Majority NFP200+ nerves are unmyelinated (arrowheads) in coronal pulp. The scale bar is 4 μm in A and 500 nm in B. |
 XML Download
XML Download