

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Kawasaki disease (KD; MIN #611775) was first diagnosed by the pediatric doctor Tomisaku Kawasaki in 1961.1 KD occurs most frequently in infants and children younger than five, and the most common symptom is continued fever. The diagnostic criteria for KD are four of the following symptoms: bilateral bulbar conjunctival injection, changes in the mucosa of the oropharynx, cervical lymphadenopathy, polymorphous rash, and changes of the peripheral extremities (e.g., erythema on the palmar and plantar regions, diffuse swelling of the hands and feet, and desquamation on the tips of fingers).2 KD is self-limited: the symptoms typically increase over two weeks and then gradually resolve on their own without treatment. Nevertheless, symptoms can be serious, presenting as a cardiac complication of coronary artery aneurysm, and therefore, intravenous immunoglobulin treatment is suggested to prevent them. The cause of KD remains unknown, although medical scientists speculate that there is a genetic influence on disease susceptibility. KD patients are increasing in Northeast Asian populations, such as Japan and Korea,3 and Taiwan has the third highest incidence rate of KD. The Asian incidence rate is more than 10-times higher than that in Caucasian populations in Europe.4
Through genome-wide association studies (GWAS), an advanced procedure has been designed to identify a number of novel genetic loci associated with KD.3 Several researchers have performed GWAS in Caucasians and Asians to confirm susceptibility loci for KD.5 We used the Affymetrix Genome-Wide Human SNP array 5.0 to perform GWAS. As a result, we selected variants of the early B cell factor 2 [EBF2 (MIM 609934)] gene. EBF2 is one of the candidate genes of KD in Korea that has not been reported in any other countries. The EBF2 gene is located on chromosome 8p21.2, and is a key transcriptional regulator of brown fat cell fate. The EBF2 gene is known to be associated with Kallmann syndrome (KS).6
In this study, we investigated genetic variants of the EBF2 gene in a Korean population with KD, and examined the relationship between genetic susceptibility of KD and variations in that gene.
MATERIALS AND METHODS
Study population
We recruited 295 KD patients from medical institutions in Korea. The patients were diagnosed from January 2012 to October 2015 at Severance Children's Hospital. Incomplete KD patients were excluded from this study. The controls were 200 unrelated healthy Korean children. Written informed consent from all participants was obtained. This study was approved by the Institutional Review Board of Yonsei University College of Medicine (IRB No. 2008-0055-010).
Genomic DNA extraction and sequencing
Genomic DNA was extracted from whole blood samples with the QIAmp DNA Blood Mini Kit (QIAGEN, Hilden, Germany), and quantified using an Epoch microplate spectrophotometer (BioTek, Winooski, VT, USA).
We analyzed a total of 15 pairs of primers and whole-exome sequencing results. The EBF2 gene was amplified by polymerase chain reaction (PCR) with an exon site-specific primer designed using extracted genomic DNA as a template. Sequencing was performed using the 3730 DNA Analyzer (Applied Biosystems, Foster City, CA, USA).
Selection of tag SNPs and SNP genotyping
The 1000 Genomes database and Haploview software (version 4.2) were used to select tag single nucleotide polymorphisms (SNPs). Five tag SNPs in the EBF2 gene (rs561367201, rs10866845, rs75171102, rs573622423, and rs901176) were selected in our study based on an r2 threshold of 0.8 and a minor allele frequency threshold of 0.01. Genetic variants of the EBF2 gene were genotyped using TaqMan® assays (Applied Biosystems). The accuracy of the results was confirmed by Sanger sequencing methods (Supplementary Fig. 1, only online). Genotyping of all samples was duplicated. The real-time PCR reaction was conducted in a final volume of 10 µL, including 15 ng of genomic DNA, 5 µL of TaqMan® Universal PCR Master Mix, and 0.25 µL of 40× TaqMan® assay. Thermal cycling conditions were as follows: initial denaturing at 95℃ for 10 min, 45 cycles of 95℃ for 15 s, and 60°C for 1 min. Genotyping was performed on a QuantStudio™ 6 Flex Real-Time PCR System (Applied Biosystems). The QuantStudio™ 6 Flex Real-Time PCR software ver. 1.2 was used for allelic discrimination.
Statistical analysis
All statistical analyses were performed using R software, version 3.4.0, on a Windows 10 platform. Allele and genotype frequencies of polymorphisms between KD patients and controls were compared using the chi-square test. Characteristics of KD patients and controls were compared using Student's t-test. Odds ratio (OR) and 95% confidence interval (CI) were calculated to explain the gene-gene interactions using binary logistic regression. A p value<0.05 was considered statistically significant.
RESULTS
Clinical features of the study population
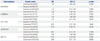
The patients with KD and healthy controls were recruited form Yonsei Medical Center in Korea. A total of 495 volunteers were enrolled in this study, including KD patients and controls (Table 1). Of the 295 patients with KD, 101 were female (34.2%), and 194 were male (65.8%). Among the control group, there were 76 males (38.0%) and 124 females (62.0%) (Table 1).
KD patients showed symptoms of fever (99.3%) and elevated white blood cell (WBCs) counts in comparison with the control group. Clinical levels of C-reactive protein (CRP), erythrocyte sedimentation rate (ESR), and lactate dehydrogenase (LDH) were increased, compared to the control subjects (Table 1). The following clinical symptoms were observed in KD patients: red-eye was found in 216 of 271 patients (79.7%), rashes in 204 of 270 patients (75.6%), and BCG injection site erythema in 71 of 268 patients (26.5%).
Polymorphisms of the EBF2 gene associated with clinical features of KD
As a result of preliminary exon sequencing of 48 chromosomes, we discovered 12 SNPs in the EBF2 gene. Among the discovered SNPs, there were no novel markers. Tagging SNPs of the EBF2 gene was selected through linkage disequilibrium (LD) analyses using the Haploview program. Therefore, five tag SNPs of the EBF2 gene were selected (Fig. 1). The genes were located at the chromosome 8p21.2 locus. Genotype and allele frequencies for these SNPs are listed in Table 2. There was a significant difference in allele frequency between KD patients and controls only for rs10866485 in the EBF2 gene (p=0.040), while a difference in genotype frequency was not observed any other markers.
We observed relationships between clinical data and selected SNPs of the EBF2 gene. The GG genotype of the rs561367201 polymorphism showed a significant association with increased WBC (p<0.000), ESR (p<0.000), CRP (p<0.000) level, and LDH (p<0.000) level. The rs10866845 polymorphism was associated with LDH (Table 3). Associations of the rs901176 polymorphism in the EBF2 gene with ESR and CRP level were confirmed (Tables 4 and 5).
Association of genetic variants in the EBF2 gene with KD
Logistic regression analysis indicated that rs10866845 and rs901176 polymorphisms were significantly associated with KD. For the rs10866845 polymorphism, the OR of C carriers (TC+CC) for the risk of KD was 1.58 (95% CI=1.06–2.34, p=0.0217) under dominant mode inheritance. The rs901176 polymorphism was associated with KD in a dominant model and recessive model (Table 6). The rs561367201 polymorphism was associated with red-eye symptoms (OR=0.25, 95% CI=0.08–0.80, p=0.0320), and the rs75171102 polymorphism was significantly related to red-lips feature (OR=1.40, 95% CI=0.33–5.95, p=0.0359) in KD patients. We did not discover any other significant ORs between KD-related clinical features and polymorphisms of the EBF2 gene (Supplementary Table 1, only online).
DISCUSSION
To date, three EBF genes have been isolated, EBF1, EBF2, and EBF3.2 EBF has a helix-loop-helix (HLH) and highly conserved transcription factors. The EBF gene is also referred to as the Collier/Olf/EBF (COE) gene.78910 COE proteins are involved in nervous and immune system development.7 EBF1 produces a family of EBF proteins expressed in B lymphocytes, osteoblasts, and adipocytes. EBF1 is an essential gene for B-cell formation, as transgenic mice with a non-functioning EBF1 gene do not produce activated B cells and immunoglobulins.11 Another family member of the EBF gene, EBF2 is not expressed in B lymphocytes, but is expressed in osteoblasts, adipocytes, and neurons.121314 The EBF2 gene has been shown to contribute to neural development and function in analysis of EBF2 knockout mice.15 The EBF2 gene has also been shown to be related to disorders affecting the peripheral nervous system, such as Charcot-Marie-Tooth disease.16
In a recent study, the EBF2 gene was shown to be associated with white adipose browning through binding with an inhibitor of differentiation 1 (Id1) known to interact with the HLH structure protein.17 EBF2 is expressed more in brown adipocytes than white adipocytes. By regulating Peroxisome Proliferator activated receptor gamma (Pparγ) binding activity,12 EBF2 helps convert white adipocytes to brown-adipocytes that consume energy by generating heat when the body is exposed to low temperatures. In contrast, abnormal accumulation of triglycerides (white adipocytes) can cause cardiac diseases or diabetes.17 Previous mouse studies have demonstrated that EBF2 is required for brown adipose tissue development. Brown adipocytes have been shown to reduce metabolic diseases and obesity in mice and rats, and a human study confirmed a correlation between weight and brown adipocytes.18
Additionally, EBF2 is a regulator of osteoclast differentiation by controlling osteoblast-dependent differentiation. When the EBF2 gene is not functional, bone mass is reduced. These functions of the EBF2 gene were demonstrated in EBF2-null mouse experiments.19 However, the EBF2 gene is not known to correlate with KD. An association of KD with genetic variants of EBF2 was not found in any population. Through genomic research, we examined the association between EBF2 gene polymorphism and KD in a Korean population.
Many inflammatory markers are generally elevated in the peripheral blood of KD patients at an acute phase. Our study confirmed that inflammatory markers, such as WBC, platelet, ESR, CRP, and LDH, were elevated in KD patients. No previous studies have reported significant associations between the EBF2 gene and inflammation markers. In our study, EBF2 gene polymorphisms were significantly associated with the inflammation markers ESR, CRP, and LDH (Tables 3, 4, and 5).
Among several symptoms of KD, cardiovascular complications are found in less than 20% of KD patients. In an analysis of associations of coronary artery complications with EBF2 gene polymorphisms, no significant correlation was observed. Additionally, KD patients with the rs573622423 variant of the EBF2 gene showed a weak association with rash symptoms (OR=0.25, 95% CI=0.06–1.00, p=0.0630).
In this study, we observed associations between KD and EBF2 gene polymorphisms. With the rs10866845 variant, the C allele was a risk factor of KD. Also, we found the rs901176 polymorphism to be a significant risk variant of KD.
In conclusion, our study of associations between genetic variants in the EBF2 gene and KD was the first in a Korean population. According to the results of our study, EBF2 is an interesting candidate gene for research on KD.
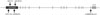





 XML Download
XML Download