

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
The proliferation of vascular smooth muscle cells (VSMCs) under pathological conditions directly contributes to the progression of cardiovascular diseases, such as atherosclerosis, hypertension, and restenosis. Among the variety of stimulants that induce VSMC proliferation, native low-density lipoprotein (nLDL) is believed to be a major independent risk factor for the development of atherosclerosis as a mitogenic molecule in lesions in which endothelial dysfunction occurs.12 In a previous study, we reported that nLDL, as a stimulant inducing VSMC proliferation, independently activates extracellular signal-regulated kinase (ERK) 1/2 mitogen-activated protein kinase (MAPK) and protein kinase C (PKC) βII. The activation of these kinases has been shown to be associated with NADPH oxidase activation through p47phox translocation to p22phox and subsequent superoxide generation, which contributes to VSMC proliferation.3
Limonin is a colorless tetracyclic triterpenoid and one of the most common bioactive compounds in citrus fruits. Limonin possesses various biological activities, including inhibition of cancer cell proliferation through induction of apoptosis4 and inhibition of aberrant crypt formation via suppression of cyclooxygenase 2 and inducible nitric oxide synthase.5 In vivo animal tests showed that limonin can inhibit 7,12-dimethylbenz[a] anthracene-induced neoplasia6 and reduce the occurrence of azoxymethane-induced colon carcinogenesis.7 In the cardiovascular system, limonin suppresses tumor necrosis factor-α-dependent p38 MAPK phosphorylation in human aortic smooth muscle cells.8 However, the mechanisms through which limonin regulates VSMC proliferation have not yet been elucidated.
Arginases catalyze L-ornithine formation via the substrate L-arginine, and the produced L-ornithine is used as a precursor for the synthesis of polyamines, spermidine, spermine, and putrescine. Increased arginase activity is involved in VSMC proliferation in a polyamine-dependent manner9 and in neointima formation in the carotid artery.10 Conversely, systematic arginase inhibition with chemical inhibitors reduces wall thickness in atherosclerotic mice.11 However, the exact mechanisms through which arginase inhibition affects VSMC proliferation under hyperlipidemic conditions remain elusive.
During a screening assay for identification of arginase inhibitors from natural medicinal products, we found that limonin inhibited several arginase isoforms. Therefore, in this study, we tested whether limonin-dependent arginase inhibition may regulate nLDL-stimulated VSMC proliferation and examined the related mechanisms.
MATERIALS AND METHODS
Materials
Limonin (batch no. 77/090915) was purchased from Nature Standard (Shanghai, China): the purity was more than 98% by high-performance liquid chromatography (HPLC). Anti-serum against p21Waf1/Cip1, p27Kip1, actin, arginase I, and arginase II were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). Phospho-PKCβII and phospho-ERK1/2 antiserum were from Cell Signaling Technology (Danvers, MA, USA). All reagents were purchased from Sigma (St. Louis, MO, USA) unless otherwise stated.
Isolation of rat aortic smooth muscle cells (VSMCs)
VSMCs were isolated from the thoracic and upper parts of the abdominal aorta in 10- to 12-week-old male Sprague-Dawley rats as previously described,12 with modifications. Briefly, stripped aortas were prepared from anesthetized rats, cut into 2-mm pieces, treated with type-II collagenase (1 mg/mL; Gibco, Waltham, MA, USA) for 1 h to remove endothelial cells, and washed with culture medium [Dulbecco's modified Eagle medium (DMEM) containing 10% fetal bovine serum, penicillin (100 U/mL), and streptomycin (100 µg/mL)]. The de-endothelialized aortic pieces were incubated with culture medium on gelatin (0.1%)-coated culture dishes for approximately 10 days. VSMCs were identified by their spindle-shaped pattern and further confirmed by double staining using platelet endothelial cell adhesion molecule-1, a specific marker for endothelial cells, and α-smooth muscle actin, a specific marker for SMCs. All cells were stained with anti-α-smooth muscle actin antibodies. The VSMCs were routinely used between the fourth and seventh passages. For all experiments, confluent cells were maintained with medium (DMEM plus 0.1% fetal bovine serum, 100 U/mL penicillin, and 100 µg/mL streptomycin) for 24 h.
nLDL preparation
nLDL (density 1.019–1.063 g/mL) was isolated from the plasma of normocholesterolemic subjects (serum cholesterol <6.2 mM) by differential ultracentrifugation as previously described.13 During LDL manipulation and storage, caution was taken to ensure nLDL integrity and prevent LDL from being oxidized. Endotoxin contents were below the detection limit (1 ng/mL), as measured using an endotoxin assay kit (Pharmingen, San Diego, CA, USA). Commercial LDL and oxidized LDL prepared by the CuSO4 reaction were purchased from Intracel (Germentown, MD, USA). Prepared LDLs from three independent experiments did not show any oxidation (−0.60±0.06 µM) in thiobarbituric acid reactive substances assays using malondialdehyde as a standard. This value was comparable to commercially available nLDL, which was −0.55±0.04 µM. That of oxidized LDL was 7.95±0.60 µM. In agarose gel electrophoresis, prepared LDL was not oxidized because the mobility of prepared LDL was significantly slower than that of oxidized LDL.
Cell proliferation assay
Cells (1×104 cells/well) were incubated in 96-well plates, starved, and stimulated with nLDL in the presence or absence of inhibitor for 24 h. Cell proliferation was measured using WST-1 reagent (Roche Co., Indiana, IN, USA) and a BrdU cell proliferation assay kit (ThermoFisher Scientific, Waltham, MA, USA) according to the suppliers' protocols.
Western blot analysis
Cell lysates prepared in sodium dodecyl sulfate-sample buffer were separated using sodium dodecyl sulfate-polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes (Bio-Rad, Hercules, CA, USA), and analyzed with primary and secondary antibodies according to the supplier's protocols. Protein bands were visualized using a chemiluminescence kit.
Arginase activity measurement
Arginase activity was measured by quantifying the amount of urea generated from L-arginine using previously described methods.14
Membrane fractionation
Treated cells were homogenized twice in Tris buffer [25 mM, pH 7.4, protease inhibitors (Roche Co.), 250 mM NaCl, 0.1% β-mercaptoethanol, and 3 mM ethylenediaminetetraacetic acid] for 3 min and centrifuged at 1000×g for 10 min to remove cell debris and unbroken cells. The supernatants were centrifuged at 21000×g for 45 min at 4℃. The cytosolic and membrane fractions containing 20 µg proteins were used for western blot analysis to assess the translocation of p47phox from the cytosol to the membrane.
Measurement of superoxide production using lucigenin
Serum-starved cells were harvested by centrifugation (800×g, 5 min) after trypsin treatment, and cells (1×105 cells) were then incubated for 30 min with various inhibitors. Lucigenin (5 µM)-induced chemiluminescence was recorded using a luminometer (Lumat LB9507; Berthold Technol., Oak Ridge, TN, USA) in Krebs-HEPES (99.01 mM NaCl, 4.69 mM KCl, 1.87 mM CaCl2, 1.2 mM MgSO4, 20 mM NaHEPES, 1.03 mM K2HPO4, 25 mM NaHCO3, 11.1 mM D-[+] glucose, pH 7.4) buffer with or without nLDL stimulation (100 µg/mL). The chemiluminescence intensity in all experiments was measured for 5 min, and the values are presented as relative lucigenin units/min.3
Measurement of mitochondrial reactive oxygen species in living cells
Cells were seeded on a glass bottom fluoro-dish coated with 0.1% gelatin, and cultured in growth or starvation media for 1 day each. MitoSox-Red (100 nM, 37℃, 30 min; Molecular Probes, Eugene, OR, USA) was used for mitochondrial superoxide measurements. Fluorescent images were acquired using an LSM780 NLO confocal microscope (Carl Zeiss, Oberkochen, Germany) with excitation/emission wavelengths of 510/580 nm. HEPES-buffered Hanks balanced salt solution was used as the imaging buffer.15 All acquired LSM raw data obtained from imaging were processed using ZEN 2011 software (Carl Zeiss).
HPLC analysis for L-arginine and polyamine quantification
The intracellular concentration of L-arginine was determined by HPLC using pre-column derivatization with o-phthalaldehyde (OPA) according to a previously published method,16 with modifications. L-arginine (100 µM) was added to cell lysate (0.1 mM) as an internal standard. The samples were extracted on solid-phase extraction cartridges (CBA Bond Elute, Varian, Palo Alto, CA, USA). The recovery rate was 87.5±3.9%. Eluates were dried over nitrogen and resuspended in double-distilled water for HPLC analysis. HPLC was performed on a computer-controlled Waters chromatography system (M600E) consisting of an automatic injector (M7725i; Waters Co., Milford, MA, USA) and a fluorescence detector (FP-1520; Jasco Co., Portland, OR, USA). Samples were incubated for exactly 1 min with OPA reagent (5.4 mg/mL OPA in borate buffer, pH 8.4, containing 0.4% 2-mercaptoethanol) before automatic injection into the HPLC. The OPA derivatives of L-arginine and polyamine were separated on a 150×4.6 mm-5 µm Zorbax Eclipse XDB-C18 column (Watertown, MA, USA), with the fluorescence detector set at an excitation wavelength of 340 nm and an emission wavelength of 450 nm. Samples were eluted from the column with 0.96% citric acid/methanol (70:30; pH 6.8) at a flow rate of 1.5 mL/min.
RESULTS
The arginase inhibitor limonin suppresses the proliferation of nLDL-stimulated VSMCs
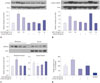
To determine the inhibitory effects of limonin (Fig. 1A), we assayed arginase II activity in the presence of different L-arginine (5–100 µM) and limonin (25 µM and 50 µM) concentrations. As shown in Fig. 1B, limonin was an uncompetitive inhibitor with the same slope (slope=0.02) and a different Y-intercept in Lineweaver-Burk plots. Next, we measured the inhibitory activity of limonin toward arginase I and II prepared from liver and kidney lysates because arginase I and II were dominantly expressed in the liver and kidney, respectively. As shown in Fig. 1C and D, limonin inhibited arginase I and II activity in a concentration-dependent manner. The maximal inhibitory activities of limonin at 50 µg/mL were 42% and 36%, and the half-maximal inhibitory concentration values were 89.33 µM and 81.65 µM for arginase I and II, respectively. Interestingly, limonin completely inhibited nLDL-induced VSMC proliferation at a concentration of 25 µg/mL in both WST-1 and BrdU assays (Fig. 1E and F). The effects of limonin on VSMC proliferation were further tested by measuring the protein expression of p21Waf1/Cip1, which regulates progression through the G1/S transition checkpoint in cell division. nLDL induced a significant decrease in the protein level of p21Waf1/Cip1, whereas limonin incubation recovered p21Waf1/Cip1 protein levels without affecting p27Kip1 (Fig. 1G). Limonin, however, had no effect on the expression of arginase I and II protein (Fig. 1H).
Limonin blocks nLDL-dependent NADPH oxidase activation through inhibition of PKCβII
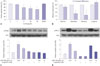
We previously demonstrated that nLDL induced VSMC proliferation through activation of NADPH oxidase and that phosphorylation of both PKCβ and ERK1/2 was responsible for p47phox translocation to the membrane.3 Therefore, we tested whether limonin prevented nLDL-induced activation of PKCβII and ERK1/2. Interestingly, limonin abolished PKCβII phosphorylation at 10 min in nLDL-stimulated VSMCs (Fig. 2A), but had no effect on the activation of ERK1/2 (Fig. 2B). Furthermore, limonin suppressed p47phox translocation into the membrane (Fig. 2C), and this effect was associated with the prevention of nLDL-stimulated superoxide generation (Fig. 2D).
Increased mitochondrial reactive oxygen species induces PKCβII phosphorylation, which is blocked by limonin treatment
To elucidate the mechanisms through which limonin inhibited PKCβII phosphorylation, we measured mitochondrial reactive oxygen species (ROS) using MitoSOX-Red. nLDL stimulation induced a significant increase in fluorescence intensity, compared with the untreated sample (nLDL vs. untreated, 25.8±1.5 vs. 14.6±0.8 AU, p<0.01). This difference was abolished following incubation with limonin incubation (Limonin+nLDL vs. nLDL, 16.1±2.3 vs. 25.8±1.5 AU, p<0.01) (Fig. 3A). Next, we tested whether increased mitochondrial ROS was involved in PKCβII phosphorylation. As shown in Fig. 3B, Mito-TEMPO, a mitochondrial ROS scavenger, inhibited nLDL-induced PKCβII phosphorylation (nLDL vs. untreated, p<0.01; Mito-TEMPO+nLDL vs. nLDL, p<0.01).
Limonin-dependent arginase inhibition increases intracellular L-arginine levels but decreases polyamine concentrations
We next analyzed the concentration of the substrate L-arginine and the polyamine products (spermine, spermidine, and putrescine) of arginase following limonin treatment. As shown in Fig. 4A, incubation of VSMCs with limonin inhibited arginase activity in a concentration-dependent manner. Limonin treatment reduced the intracellular polyamine concentrations to 85.6±9.2% for spermine, 41.6±15.9% for spermidine, and 31.1±26.5% for putrescine. However, L-arginine concentrations were significantly increased to 116.5±2.8% (Fig. 4B). In untreated VSMCs, the concentrations of intracellular L-arginine, spermine, spermidine, and putrescine were 248.2, 113, 11.7, and 3.4 µM, respectively. Intracellular L-arginine concentration was consistent with those in a previous study (100–800 µM) in cultured cells.17181920 Based on this result, we next tested whether increased the L-arginine content would affect the activation of PKCβII and ERK1/2. Interestingly, pre-incubation with L-arginine prevented nLDL-dependent PKCβII activation as early as 10 min (Fig. 4C), although L-arginine had no effect on nLDL-induced ERK1/2 activation (Fig. 4D). These results implied that the limonin-dependent increase in L-arginine concentrations inhibited nLDL-induced NADPH oxidase activation, which was associated with VSMC proliferation. Therefore, limonin may be useful for the treatment of hypercholesterolemia-dependent proliferation of VSMCs.
DISCUSSION
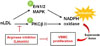
In the current study, we demonstrated that limonin is an uncompetitive inhibitor of arginase I and II isoforms, acting in a concentration-dependent manner without affecting protein levels. Increased L-arginine concentrations following treatment with limonin prevented mitochondrial ROS-dependent PKCβII activation, which is associated with NADPH oxidase activation, and then abolished nLDL-stimulated VSMC proliferation in a p21Waf1/Cip1-dependent manner (Fig. 5). Our findings suggested that limonin may have beneficial effects on prevention of VSMC proliferation in pathological conditions, such as hypertension, restenosis, and atherosclerosis.
Although ROS can act as a second messenger, influencing discrete signal transduction pathways, such as maintenance of differentiated and contractile phenotypes, under normal physiological conditions, oxidative stress is known to serve an important role in hyperlipidemic vascular injury (e.g., hypertrophy). NADPH oxidases are multiprotein complexes containing two membrane-bound subunits (p22phox and Nox) and potentially three cytosolic subunits (i.e., Rac1, p47phox, and p67phox), which are recruited upon activation to the membrane-bound Nox/p22phox complex. Many stimuli activate p47phox-regulated NADPH oxidase in VSMCs.21222324 Angiotensin II induces the release of arachidonic acid by PLA2, resulting in activation of NADPH oxidase.23 Additionally, lysophosphatidylcholine treatment triggers rapid translocation of p47phox to the membrane, thereby increasing NADPH oxidase activity and growth.24 The phosphorylation of p47phox is essential for its translocation and NADPH oxidase activation. The kinases responsible for p47phox activation are PKC and ERK1/2, which can be phosphorylated at Ser359/370/379 and Ser345, respectively.252627 In addition, we recently demonstrated that nLDL induces activation of NADPH oxidase through p47phox translocation to p22phox by independently phosphorylating PKCβII and ERK1/2. Furthermore, generation of the superoxide anion from activated NADPH oxidase 1 plays a critical role in VSMC proliferation.3 In this study, we found that limonin acts as an uncompetitive arginase inhibitor and prevents nLDL-induced PKCβII phosphorylation without affecting ERK1/2 phosphorylation, suppressing p47phox translocation to the membrane and subsequently blocking ROS production. Limonin effectively prevented nLDL-stimulated VSMC proliferation in a p21Waf1/Cip1-dependent manner. Members of the PKC family have been commonly implicated in regulation of cell cycle progression and differentiation. PKC plays critical role as an important regulator of critical cell cycle transitions, including cell cycle entry and exit and the G1/G2 checkpoints. PKC-dependent regulation of these transitions can be negative or positive, depending on the timing of PKC activation over the cell cycle and on the specific PKC isozymes involved. Most of the information relates to the participation of this PKC family in negative regulation of these transitions. A major target for PKC-dependent inhibition of cell cycle progression is the Cip/Kip cyclin-dependent kinase (cdk) inhibitor p21waf1/cip1. Increased expression of p21waf1/cip1 blocks cdk2 activity in the G1 phase. p21waf1/cip1 is a major target of p53 transcriptional activity and thus is associated with linking DNA damage to cell cycle arrest. PKC signaling can also trigger a coordinated program of pocket protein regulation leading to cell cycle withdrawal into G0.
Mitochondrial ROS is an important product of cell metabolism in the mitochondrial respiratory chain, and is associated with the development of hypertension through enhanced NADPH oxidase activity and reduced nitric oxide bioavailability. Furthermore, MitoTEMPO, a ROS scavenger, restores the deleterious effects of mitochondrial ROS.28 Interestingly, in our study, nLDL induced an increase in mitochondrial ROS production, which was abolished by limonin incubation. The elimination of increased mitochondrial ROS using MitoTEMPO blocked nLDL-induced PKCβII phosphorylation. This result was consistent with a previous report showing that inhibition of mitochondrial ROS by MitoTEMPO suppressed AdPKCβII-induced endothelial dysfunction.29
Furthermore, limonin-dependent inhibition of arginase increased intracellular L-arginine concentrations, whereas the concentrations of the polyamines spermine, spermidine, and putrescine were decreased in VSMCs. The increased L-arginine contributed to prevention of nLDL-induced PKCβII phosphorylation without affecting ERK1/2 phosphorylation, consistent with the effects of limonin. Thus, our findings, together with the effects of L-arginine on peroxynitrite-induced endothelial dysfunction,30 supported that L-arginine affected nLDL-induced VSMC proliferation. Therefore, increased L-arginine bioavailability by arginase inhibition may have beneficial effects on vascular cells, such as endothelial and smooth muscle cells, under pathological conditions. Although limonin showed beneficial effects on nLDL-stimulated VSMCs proliferation, its application in patients is limited, as effects have only been studied in animal cells, and we did not confirm the effect to many mitogens exerting VSMC proliferation.
Two isoforms of arginase (i.e., arginase I as the hepatic isoform and arginase II as the extrahepatic isoform) are encoded by distinct genes. Although arginases expression in VSMCs is constitutive and at low levels, their expression is further induced by various stimuli, such as interleukin (IL)-4, IL-13,31 transforming growth factor-β,32 lysophosphatidylcholine,33 and mechanical strain.34 Increased arginase and ornithine carboxylase activities also augment polyamine, spermine, spermidine, and putrescine, synthesis, thereby contributing to VSMCs proliferation.12 Combined with previous demonstrations representing a cGMP-dependent mechanism,35 our results suggests that arginase inhibition suppresses nLDL-induced VSMC proliferation in an NADPH oxidase-dependent mechanism.
In summary, we found that increased intracellular L-arginine by arginase inhibition had beneficial effects on the prevention of VSMC proliferation in the presence of high concentrations of nLDL. However, it is still unclear whether L-arginine supplementation may have the same effect. Indeed, clinical studies in oral L-arginine therapy in vascular diseases have led to controversial results.363738 Notably, increased arginase activity and enzymatic products (e.g., L-ornithine and polyamines) have been shown to be involved in neointima formation10 and VSMC proliferation.9 Accordingly, limonin, as an uncompetitive arginase inhibitor, may be useful for the treatment of vascular diseases associated with VSMC proliferation and hypercholesterolemia.


 XML Download
XML Download