

 PDF
PDF ePub
ePub Citation
Citation Print
Print



Antifreeze proteins (AFPs) are diverse proteins that help the survival of organisms in subzero temperatures. These proteins work in a non-colligative manner and permit the organisms to survive by lowering the freezing point below the melting point, which is known as thermal hysteresis.1 They adsorb to ice surfaces, bind to ice planes, and prevent the growth of ice crystals until temperatures reach the freezing point. With these features, AFPs have been shown to inhibit ice recrystallization (IR).2
Previous studies have demonstrated the beneficial effects of AFP on mouse ovaries, and treatment with AFPs has been used during both the vitrification and warming procedures.34 However, previous studies have found that IR inhibition during warming seems more crucial than ice crystallization inhibition during freezing.56 The aim of this study was to compare a new AFP treatment protocol in which AFP treatment is used only during the warming procedure and the conventional AFP treatment protocol in which AFP treatment is used during both the vitrification and warming procedures.
Five-week-old B6D2F1 female mice (Orient Co., Seongnam, Korea) were housed under a 12-h light/dark cycle at 22℃ and provided food and water ad libitum. This study was performed with the approval of the Institutional Animal Care and Use Committee of Seoul National University Bundang Hospital (BA1707-227/061-01). Vitrification-warming for mouse ovaries was performed by optimized protocols, as previously described.7 We added 10 mg/mL of LeIBP into the vitrification and/or warming solutions according to our previous study demonstrating the optimal AFP type and concentration for the vitrification-warming process of the mouse ovary. We designed three groups for this experiment: 1) vitrified-warmed control; 2) LeIBP-all group, LeIBP supplementation was used in two-step vitrification solutions and the first step of the warming solution; and 3) LeIBP-w group, LeIBP was added only to the first step of the warming solution.
A total of 42 ovaries obtained from the B6D2F1 mice (vitrified-warmed: n=12, LeIBP-all: n=14, and LeIBP-w: n=16) were randomly allocated into the three groups. The ovaries embedded in paraffin blocks were serially sectioned at a thickness of 5 µm. For slide staining, five slides at 100-µm intervals (5 slides for each ovary) were chosen and stained with hematoxylin (DAKO, Seoul, Korea) and eosin (Merck, Darmstadt, Germany) (H&E), terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling (TUNEL) assay, or γH2AX/Rad51 immunohistochemistry. The stained slides were read by a single experienced researcher (HSK).
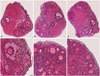
The H&E-stained ovaries were read to evaluate follicle developmental stages and morphological integrity using a light microscope (Nikon, Tokyo, Japan). The follicle developmental stages were classified in accordance with the following categories8: 1) primordial follicles, a single layer of flattened pregranulosa cells; 2) primary follicles, a single layer of cuboidal granulosa cells; 3) secondary follicles, two or more layers of cuboidal granulosa cells, with the antrum absent; and 4) antral follicles, multiple layers of cuboidal granulosa cells, with the antrum present. The primary, secondary, and antral follicles were categorized as growing follicles. The follicles were considered to not be intact if they had pyknotic bodies within granulosa cells, condensed oocyte nuclei, shrunken oocytes, oocyte cytoplasm vacuolization, or low cellular density.9 Otherwise, the follicles were categorized as morphologically intact follicles.
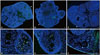
Follicle apoptosis was evaluated by an In Situ Cell Death Detection Kit (Roche, Basel, Switzerland), as previously described.10 Ovaries were then mounted with VECTASHIELD® Mounting Medium with 4′,6-diamidino-2-phenylindole (Vector Laboratories, Burlingame, CA, USA), and examined under an inverted Zeiss AX10 fluorescence microscope (Carl Zeiss, Oberkochen, Germany). The cells that had fragmented DNA displayed green fluorescence, while intact normal cells showed blue fluorescence. Follicles with over 30% of apoptosis-positive cells or oocyte nucleus were considered as apoptotic follicles.371011
Immunohistochemical analysis for DNA double-strand (DDS) breakdown and repair was performed using γH2AX antibody (1:100; Millipore, Burlington, MA, USA) and Rad51 antibody (1:100; Bioworld Technology, St. Louis Park, MN, USA), respectively.3 For secondary antibody, Alexa 594 (1:1000; Life Technology, Carlsbad, CA, USA) antibody was used and counterstained with VECTASHIELD® Mounting Medium with 4′,6-diamidino-2-phenylindole. All slides were examined under an inverted Zeiss AX10 microscope. Follicles which contained at least one nucleus stained with γH2AX were considered as follicles with DDS breakdown. In the same way, follicles containing at least one nucleus stained with Rad51 were regarded as follicles with DNA repair.3
The chi-square test was performed by SPSS software, version 12.0 (SPSS Inc., Chicago, IL, USA) to analyze the effect of each LeIBP supplementation on follicular integrity, apoptosis, and DDS breakdown/repair ratios. Results were considered significant when the probability showed a p-value <0.05.
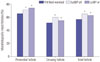
The representative images of H&E stained-ovaries are shown in Fig. 1. A total of 5558 follicles were evaluated for morphologically intact follicle ratios in accordance with the follicle developmental classification (vitrified-warmed: 1538, LeIBP-all: 1913, and LeIBP-w: 2107). The treatment with LeIBP significantly improved the integrity of primordial and growing follicles in the LeIBP-all and LeIBP-w groups, when compared with the vitrified-warmed group (vitrified-warmed, LeIBP-all, and LeIBP-w groups: 66.6, 73.0, and 74.4% for primordial follicles, and 50.2, 60.3, and 55.3% for growing follicles, respectively). Between the two LeIBP-treated groups, the LeIBP-all group showed a higher intact growing follicle ratio than the LeIBP-w group; however, the intact primordial follicle ratio was similar between the two groups (Fig. 2).
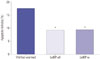
Representative images of TUNEL-stained ovaries are shown in Fig. 3. Compared to the vitrified-warmed group, the apoptotic follicle ratios of the two LeIBP treated groups were significantly decreased (vitrified-warmed 17.5%, LeIBP-all 9.2%, LeIBP-w 9.3%). No significant difference was found between the LeIBP-all and LeIBP-w groups (Fig. 4).
Immunohistochemical images of γH2AX and Rad51 stained ovaries are shown in Fig. 5. The percentage of γH2AX-positive follicles were significantly lower in the LeIBP treated groups (vitrified-warmed: 36.9%, LeIBP-all: 31.2%, LeIBP-w: 29.6%). The ratios of Rad51-positive follicles were also significantly lower in the LeIBP-all (31.6%) and LeIBP-w (32.8%) groups in comparison to the vitrified-warmed group (38.4%) (Fig. 6).
Ice formation in cells and tissues during cryopreservation is critical to cell/tissue survival as many studies have already emphasized. However, the warming process and rates also have a powerful influence on whether the cell/tissue survives or not.612 During the warming process of ovarian tissue, IR can cause rupture of the cell membrane and cell dehydration inducing lethal damage in the tissue.13 Because AFPs can prevent ice-crystallization during vitrification and recrystallization during warming, AFP treatment is known to increase survival after cryopreservation.314
A previous study demonstrated the cryoprotective effect of three different types of AFPs for mouse ovary vitrification-warming.3 Among the different types of AFPs, 10 mg/mL of LeIBP was the most effective AFP concentration when it was used for treatment during both vitrification and warming. However, there has not been any study in which AFP treatment was used during only the warming procedure, particularly the first step of the warming procedure (this step includes a very large temperature change for the ovary) to inhibit IR and improve the ovary-warming conditions. Thus, the new LeIBP protocol (LeIBP-w group) with 10 mg/mL of LeIBP treatment during the first-step warming procedure was performed in this study, and it was compared with the conventional LeIBP protocol (LeIBP-all group), which added LeIBP to the vitrification and first-step warming procedures, to find the optimal LeIBP treatment protocol in mouse ovary vitrification-warming.
In the present study, intact follicle ratios were significantly increased in LeIBP-treated groups, compared to the vitrified-warmed group, regardless of follicle developmental stage. From the results of the LeIBP-all group, we confirmed the conventional LeIBP treatment protocol was effective to preserve follicle morphology after mouse ovary vitrification-warming, which was consistent with the results of our previous studies showing its reproducibility.3 Additionally, the LeIBP-w group showed not only a significantly higher ratio of intact follicles than the vitrified-warmed group but also similar results to the LeIBP-all group. Between the two LeIBP-treated groups, only the ratio of intact growing follicles in the LeIBP-all group was significantly higher than that in the LeIBP-w group. Growing follicles are more vulnerable to cryodamage than primordial follicles due to their large size. Thus, it seems the LeIBP treatment during vitrification in the LeIBP-all group showed more powerful antifreeze effects on growing follicles than primordial follicles, and made a difference between the LeIBP-treated groups. However, primordial and total follicles showed similar results between the LeIBP-treated groups. Therefore, we suggest the new LeIBP treatment protocol can effectively preserve follicle morphology, even if LeIBP was used during only in the first step of warming. In the previous study, which compared different types of AFPs, it was clearly demonstrated that the AFP with the highest IR activity, which was LeIBP, showed the best result in the vitrification-warming procedure, even though the thermal hysteresis value of the LeIBP was the lowest among the different types of AFPs.3 Therefore, we can suggest that the main cryoprotective mechanism of AFP treatment during vitrification-warming is related to IR inhibition activity.
Follicle apoptosis, which could be induced by cryodamage, was significantly decreased in both LeIBP-treated groups, compared with the vitrified-warmed. Similar to our study, treatment with different types of AFPs has been found to exert antiapoptotic effects on rat hearts and mouse oocyte and ovary preservation at subzero temperatures.341415 Between the LeIBP-all and LeIBP-w groups, no significant difference was found in terms of apoptotic follicle ratios. Based on these data, we can speculate that the new AFP treatment protocol using LeIBP during the first step of the warming procedure can reduce follicle apoptosis to the same degree as that achieved with the conventional protocol.
γH2AX is a phosphorylated histone, H2AX on Serine 139, and is found in damaged DNA with DDS.16 Previous studies have reported that mechanical and osmotic stress occurring during the cryopreservation process induced DDS breakdown, followed by severe or lethal mutations of the cells.1718 Moreover, γH2AX has been shown to have a correlation with DNA repair, in which the repair protein Rad51 is recruited to nuclear foci after DDS damage.19 To investigate DDS damage and DNA repair in follicles, γH2AX and Rad51, respectively, were immunostained in ovarian follicles. The proportion of γH2AX-positive follicles was significantly decreased in both LeIBP-treated groups, compared to the vitrified-warmed group. Similar to the γH2AX staining result, Rad51 immunostained follicle ratios were also significantly reduced in both LeIBP-treated groups, compared to the vitrified-warmed group. The outcomes of the LeIBP-all group were consistent with those in our previous studies that showed significantly reduced DDS damage/repair when vitrifying and warming mouse oocyte and ovary with the LeIBP treatment.314 Moreover, in the present study, the LeIBP-w group showed similar results to the LeIBP-all group with regard to the γH2AX and Rad51 immuno-stained follicle ratios. Thus, we demonstrated that the new LeIBP treatment protocol has a beneficial effect in terms of ovarian follicle DNA protection, the same as that with the conventional LeIBP treatment protocol.
Compared to the conventional vitrification-warming protocol with LeIBP-treatment (LeIBP is added in both vitrification and warming solutions), we obtained comparable cryoprotective results from the new vitrification-warming protocol with LeIBP treatment in which the LeIBP treatment was used during only the first-step warming protocol. With the new LeIBP treatment protocol, we can minimize not only the amount of AFPs (incurring lower costs), but also the potential side effects of these proteins, such as induced membrane leakage, due to high AFP concentration.20 Therefore, this new protocol that requires less AFP and offers lower costs can be modified and used for future applications, such as cell, tissue, and organ cryopreservation.
This is the first study to highlight the beneficial effects of AFP treatment during only the warming procedure in mouse ovary vitrification-warming. We demonstrated that the new LeIBP treatment protocol is comparable to the conventional protocol in terms of preserving follicle morphology, reducing apoptosis, and DDS damage/repair, compared with the vitrified-warmed. Based on these results, we speculate that inhibition of IR during the warming procedure is critical to the outcomes of the vitrification-warming procedure, thereby the addition of LeIBP into the warming solution only is sufficient to give cryoprotective effects. Since only microscopic evaluations were performed, further studies to investigate the exact IR inhibition mechanism of AFPs in ovarian tissue cryopreservation are required.


 XML Download
XML Download