

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
With notable sex-related differences in the lengths of QT intervals, hormonal influences on cardiac repolarization have been proposed.1 Generally, women have longer corrected QT (QTc) intervals than men, and these vary during the menstrual cycle and pregnancy, suggesting that an intrinsic hormonal regulation occurs in female hearts. Recent experiments in rabbits have demonstrated a potentially detrimental effect for estrogen on the development of cardiac arrhythmias, while progesterone was found to have a protective role.2 Therein, estrogen enhanced, while progesterone reduced, L-type calcium currents in rabbit hearts. In humans, data from long QT type 2 patients suggest that pregnancy-related hormones have protective effects against arrhythmia, whereas the postpartum period is associated with an increased susceptibility to arrhythmia.3 During late pregnancy, the maternal heart needs to adapt to significantly increased circulatory needs. Moreover, pregnancy-induced electrocardiogram disturbances are often observed, such as prolongation of the QT-interval accompanied by downregulation of Kv4.3, one of the key ion channels for repolarization.4 However, the mechanism of QT prolongation during pregnancy has not been fully elucidated.
Serotonin (5-hydroxytryptamine, 5-HT) increases during pregnancy and plays a critical role in fetal development. Serotonin affects craniofacial, gastrointestinal, and cardiovascular morphogenesis in chickens, rats, and mice.56 As sites for early serotonin biosynthesis have not been detected in embryos or extraembryonic structures,7 maternal serotonin is thought to be the major source of fetal serotonin and an important determinant for normal fetal development.8 Serotonin is primarily found in the gastrointestinal tract, blood platelets, and the central nervous system of animals, including humans. However, the main source and pathway of serotonin regulation during pregnancy has not been clarified. Moreover, the effects of highly elevated serotonin levels in the mother have not been investigated. Two Htr3 antagonists, granisetron (Kytril) and ondansetron (Zofran), induce QT prolongation at high concentrations and are associated with an elevated risk of cardiac dysrhythmias in patients.9 Recently, we reported that 5-HT3a-receptor (Htr3a) knockout mice (Htr3a−/−) succumb to sudden death, exhibiting QTc prolongation and fatal arrhythmias during pregnancy.10 Therefore, we hypothesized that serotonin would prevent further QT prolongation during pregnancy via Htr3a. To test this hypothesis, we directly measured Kv currents in freshly-isolated left ventricular (LV) myocytes from wild-type non-pregnant (WT-NP) and late-pregnant (WT-LP), as well as from non-pregnant Htr3a−/− mice (Htr3a−/−-NP), after administration of serotonin and its antagonist. We also measured Kv4.3-Htr3a interactions by co-immunoprecipitation and the membrane trafficking of Kv4.3 after serotonin administration.
MATERIALS AND METHODS
This investigation was carried out in accordance with the Guide for the Care and Use of Laboratory Animals, published by the US National Institutes of Health (NIH Publication, 8th Edition, 2011). This study protocol was approved by the Institutional Animal Care and Use Committee of the Yonsei University College of Medicine (2016-0189) and the Cardiovascular Research Institute.
Single-cell electrophysiological recordings and data analysis
For patch-clamp study, ventricular myocytes were isolated from adult (8–10 weeks of age) C57BL6 mice by perfusing Ca2+-free normal Tyrode's solution containing collagenase (1.2 mg/mL−1, Type II, Worthington Biochemical, Lakewood, NJ, USA) on a Langendorff column at 37℃ as described previously.11 Isolated ventricular myocytes were kept in high K+, low Cl− solution at 4℃ until ready for use. Ito current recordings from single isolated ventricular cardiac myocytes were performed using the whole-cell voltage clamp technique with an Axopatch-200B amplifier (Axon Instruments Inc., Foster City, CA, USA), controlled by Clampex 10 a Digidata 1550 (Axon). Data were low pass-filtered at 5 kHz and sampled at 10 kHz. The patch pipettes were made of borosilicate glass (Harvard Apparatus, Holliston, MA, USA) and pulled with a Narishige puller (PC-10; Narishige, Tokyo, Japan). All recordings were carried out at room temperature. The extracellular solution contained (mmol/L) 136 NaCl, 10 glucose, 10 HEPES, 2 MgCl2, 1 CaCl2, 4 KCl, and 5 CoCl2 (pH adjusted to 7.4 with NaOH). The intracellular recording pipette solution contained (mmol/L) 135 KCl, 10 ethylene glycol tetraacetic acid (EGTA), 10 HEPES, and 1 MgCl2 [pH adjusted to 7.2 with potassum hydroxide (KOH)]. Tetrodotoxin inhibits Na+ currents; CoCl2 inhibits Ca2+ currents. For HERG current (Ikr) recording, bath solution contained (mmol/L) 140 NaCl, 3.5 KCl, 1.5 CaCl2, 1.4 MgSO4, and 10 HEPES (pH adjusted 7.4 with NaOH). The pipette solution contained (mmol/L) 140 KCl, 1 CaCl2, 2 MgCl2, 10 HEPES, 11 EGTA, 5 Na2-ATP, and 5 creatine phosphate (disodium salt) (pH 7.2 adjusted with KOH). Serotonin, m-CPBG, ondansetron, 4-AP, or tetraethylammonium ions (TEA) were applied to isolated myocytes during the recordings, using narrow-bore capillary tubes (300 µm i.d.) placed within ≈200 µm of the cell.
The Kv currents recording protocols were also performed as previously described.11 Briefly, we used pre-pulse to −40 mV for 25 ms to in-activated INa. Kv current was recorded in the voltage-clamp mode with 4 s pulses from a holding potential of −70 mV, with different test potentials increased from −40 mV to +60 mV with 10-mV steps. The currents were normalized to the cell membrane capacitance and averaged. The values were not corrected for the junction potential (pipette offset). This was compensated prior to giga-seal formation. Additionally, we examined potassium outward currents in adult (8–10 weeks of age) SD rats and adult rabbits. The Patch clamp data were analyzed using p-Clamp software, version 10.4 (Axon Instruments) and Origin Pro, version 9.0 (Origin Lab Corp., Northampton, MA, USA). All values are provided as means±SEMs. Current densities (pA/pF) were obtained after normalization to cell surface area calculated by Patch master.
An expanded methods section is available in the Supplementary Material (only online). The authors had full access to and take full responsibility for the integrity of the data. All authors have read and agreed to the contents of the manuscript.
RESULTS
Pregnancy increases serotonin and serotonin-related molecule levels
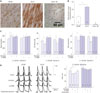
To test whether pregnancy influences Htr3a levels in the myocardium, we performed immunohistochemistry. We found that Htr3a expression was detectable in the myocardium of WT-NP and WT-LP mice, but not in that of Htr3a−/− mice (Fig. 1A). Htr3a serotonin receptor protein levels were higher in WT-LP than in WT-NP mouse hearts (2.3±0.2 vs. 1±0, respectively, p<0.001) (Fig. 1B). Serum serotonin concentrations in WT-LP mice increased almost 1000-fold, compared to that in WT-NP mice (866.7±20.9 ng/mL vs. 1±0 ng/mL, p<0.001). Serotonin levels in the ventricle was also higher in WT-LP than in WT-NP hearts. Moreover, the transcript levels of tryptophan hydroxylase 1 (Tph1), a rate-limiting enzyme of 5-HT synthesis, were higher in ventricles from WT-LP hearts than in those from WT-NP hearts (Supplementary Fig. 1, only online).
Serotonin shortens QT intervals and APD
WT-LP mice displayed more prolonged QT and QTc intervals than NP mice (p<0.001) (Supplementary Fig. 2A, only online). The action potential duration at 90% (APD90) measured at the base of the left ventricle was more prolonged in WT-LP mice than in NP mice (p<0.001) (Supplementary Fig. 2B, only online). Compared with WT-NP mice, WT-LP mice hearts were heavier (p<0.001) (Supplementary Fig. 2C, only online) and presented larger LV end-diastolic dimensions (p=0.005) (Supplementary Fig. 2D, only online).
We examined the effects of serotonin on heart rate, QT, and QTc intervals between WT-NP and WT-LP mice. Intra-peritoneal injections with 100 µmol/L of serotonin significantly shortened the QT (50.9±3.8 ms vs. 38.7±2.4 ms, p<0.001) and QTc intervals (56.5±3.2 ms vs. 45.2±3.1 ms, p<0.001) in WT-NP mice, but not in WT-LP mice (Fig. 1C). In whole hearts retrogradely perfused with serotonin at 100 nmol/L, the APD90 was shortened in both WT-NP (from 54.9±3.1 to 44.1±2.7 ms, p<0.001) and WT-LP mice (from 63.4±4.7 to 49.4±4.0 ms, p<0.001). These serotonin effects were mimicked by m-CPBG, an Htr3 agonist. When the animals were pretreated with the Htr3 antagonist ondansetron (1 µmol/L), serotonin failed to shorten APD90 values (Fig. 1D). These results indicate that serotonin treatment shortens APD and consequently QT intervals by acting on Htr3.
Effects of serotonin and an Htr3 agonist on Kv current
To clarify the molecular determinants of serotonin-induced APD shortening, we examined Kv channel properties. Whole-cell path clamp recordings were obtained from LV myocytes isolated from adult WT-NP and WT-LP mice. Fig. 2A shows the change in Kv currents by various pharmacological interventions. Kv current densities were 25% lower in WT-LP mice than in WT-NP mice (22.3±2.9 vs. 34.3±1.4 pA/pF, p<0.001). Serotonin (100 µmol/L for 10 min) and m-CPBG (30 µmol/L for 10 min) increased Kv current densities in WT-NP, but not in WT-LP mice. The co-application of ondansetron (0.5 µmol/L for 10 min) abolished the increases in Kv current densities observed in WT-NP mice following serotonin treatment.
Fig. 2B shows the current-voltage (I–V) relationships of Kv current densities in WT-NP (left panel) and WT-LP mice (right panel), respectively. Kv current densities were significantly decreased in WT-LP mice, compared to those in WT-NP mice. Serotonin increased the amplitude of peak Kv current densities (at +60 mV) in WT-NP, but not in WT-LP mice (34.3±1.4 vs. 40.5±2.3 pA/pF, p=0.002). Likewise, m-CPBG (30 µmol/L), an Htr3a agonist, increased peak Kv current densities (at +60 mV) in WT-NP, but not in WT-LP mice (36.3±1.4 vs. 41.3±1.2 pA/pF, p=0.002) (Fig. 2C).
We antagonized serotonin receptors by applying ondansetron to freshly-isolated LV myocytes from WT-NP or WT-LP mice. Even at a high concentration, ondansetron (5 µmol/L for 10 min) did not decrease Kv current densities in myocytes from WT-NP mice. Conversely, ondansetron significantly decreased the Kv current densities in myocytes from WT-LP mice, suggesting that, in the WT-LP heart, serotonin signaling is already saturated (Supplementary Fig. 3, only online).
Ikr tail currents in WT-NP and Htr3a−/−-NP adult mouse LV and LV apex myocytes were relatively small and did not change in either the WT-NP or LP mice (Supplementary Fig. 4, only online).
Serotonin activates Htr3a-mediated membrane trafficking
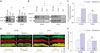
Kv4.3 protein expression was decreased more in WT-LP mice than in WT-NP mice (1.0±0.0 vs. 0.4±0.0, p<0.001). The protein expressions of Kv1.5, Kv1.4, Kv4.2, and HERG did not differ between the WT-NP and LP mice (Supplementary Fig. 5, only online). Therefore, we hypothesized that an interaction may exist between Kv4.3 and Htr3a. Fig. 3A demonstrates that Kv4.3 is directly associated with Htr3a in WT-NP and WT-LP mice. Interestingly, this interaction increased upon serotonin treatment (100 µmol/L for 1 h) in WT-NP LV myocytes, but not in WT-LP LV myocytes (Fig. 3B and C). Confocal microscopy revealed that Htr3a and Kv4.3 co-localized at the plasma membrane and in the cytoplasm following pre-treatment with serotonin (100 µmol/L for 1 h) (Fig. 3D). Upon serotonin stimulation, Kv4.3 and Htr3a were translocated to the surface of the membrane in WT-NP, but not in WT-LP mice (Fig. 3E).
Because treatment with serotonin caused the translocation of Kv4.3 and Htr3a to the plasma membrane, we performed cellular fractionation to assess the distribution of these proteins. Treatment with serotonin and the agonist increased the abundance of Kv4.3 in the membrane fraction. Conversely, the co-application of serotonin and ondansetron (0.5 µmol/L for 1 h) abolished this phenomenon in WT-NP mouse hearts (Supplementary Fig. 6A, only online). However, in WT-LP mouse hearts, the same robust abundance of Kv4.3 in the membrane fraction upon serotonin treatment was not observed (Supplementary Fig. 6B, only online).
Serotonin improves Kv4.3 trafficking through its interaction with Hsc70, but not HSP90
Enhanced plasma membrane localization of the KCNH2 channel was previously shown to be associated with increased interactions with heat-shock proteins, despite unchanged levels of chaperone or ion-channel proteins.12 Therefore, we investigated the effect of heat-shock proteins on Kv4.3 trafficking. We demonstrated that Kv4.3 directly associates with Hsc70 in WT-NP and WT-LP mice and that this interaction increases upon serotonin treatment (100 µmol/L for 1 h) in WT-NP ventricular myocytes. However, in WT-LP myocytes, serotonin treatment did not further enhance the protein-protein interactions between Kv4.3 and Hsc70 (Fig. 4A). Moreover, the Kv4.3 ion-channel protein and Hsc70 co-localized at the membrane and in the cytoplasm of WT-NP and WT-LP ventricular myocytes. Following serotonin treatment (100 µmol/L for 1 h), Kv4.3 and Hsc70 were translocated to the membrane surface in WT-NP, but not in WT-LP mice (Fig. 4B). Confocal microscopy revealed that Kv4.3 and Hsc70 co-localized at the plasma membrane and in the cytoplasm following pre-treatment with serotonin (100 µmol/L for 1 h). Upon serotonin stimulation, Kv4.3 and Hsc70 were translocated to the surface of the membrane in WT-NP, but not in WT-LP mice (Fig. 4C and D).
To further confirm the localization of Kv4.3 and Hsc70 following treatment with serotonin, we performed cellular fractionation. Serotonin and m-CPBG increased Kv4.3 protein abundance at the plasma membrane via Hsc70. The co-application of serotonin and ondansetron (0.5 µmol/L for 1 h) abolished the translocation of Kv4.3 to the membrane in WT-NP (Supplementary Fig. 7A, only online), but not in WT-LP mice (Supplementary Fig. 7B, only online).
DISCUSSION
The present translational study provides evidence that elevated serotonin levels are associated with shorter QTc intervals through the acceleration of Kv current densities in mice. Serotonin acts on Kv4.3 channels by promoting enhanced Htr3a-mediated interactions with chaperones, augmenting membrane trafficking and increasing the repolarizing current. During pregnancy, the Htr3a-mediated Kv4.3 membrane trafficking was saturated. Elevated serotonin levels counterbalanced pregnancy-related QT prolongation by facilitating Htr3-mediated Kv currents.
Hormonal modulation of cardiac repolarization
The QTc interval is longer in women than in men,1 and women have an increased risk of drug-induced QT-interval prolongation and torsade-de-pointes tachycardia.13 Systematic studies of hormonal effects (e.g., menstrual cycle) are sparse. Studies on the hormonal regulation of the QT-interval have often relied on pooled data that are potentially flawed by large inter-individual inherent variabilities in QT-intervals.14 A recent study showed that elevated estradiol levels were associated with shorter QTc intervals in healthy women and female long QT type 2 patients. Estradiol acts on KCNH2 channels via an enhanced estradiol receptor-mediated interaction with HSP90, augments membrane trafficking and increases the repolarizing current.12
Our study shows that serotonin also affects cardiac repolarization. The QT shortening effect of serotonin was mainly mediated by increased Kv current densities via Htr3. The involvement of Htr3 is also supported by the different responses to serotonin and m-CPBG in Htr3a−/− mice.
The cellular mechanism underling the Kv4.3 and Htr3a pathway
Trafficking of the KCNH2 channel is well established. This channel traffics relatively inefficiently by itself15 and depends on chaperones for proper folding16 and trafficking to the plasma membrane.17 Trafficking-deficient long QT type 2 mutants interact with Hsc70 and HSP90 when retained in the endoplasmic reticulum.16 The recovery of channel trafficking is coupled to the dissociation of channel-chaperone complexes, suggesting that the exit from the endoplasmic reticulum is linked to the dissociation of KCNH2 from this complex.16 The cardiac HERG K+ channel is important for cardiac repolarization in humans and large animals.18 However, the role of IKr in repolarization is minor in mice.19 Consistently, the IKr was small, and the effect of serotonin on the IKr was negligible in this study.
We show that the activation of Htr3a by serotonin increases the trafficking of Kv4.3 via Hsc70. Moreover, co-immunoprecipitation illustrated enhanced co-precipitation of Htr3a and Kv4.3 following serotonin stimulation in WT-NP, but not in WT-LP mice. This finding suggest a direct interaction between serotonin (Htr3a) and Kv channels. Our work suggests that an increase in chaperone/channel complexes may occur similarly in response to serotonin stimulation by enhancing channel trafficking and membrane stability via improved folding and trafficking.
The effect of serotonin on cardiac repolarization during pregnancy
Maternal serotonin is required for normal embryonic development, as revealed by the Tph1-invalidated mouse line, in which blood is depleted of serotonin.8 Maternal serotonin also influences cardiac function in adult offspring. However, the role played by an elevated serotonin level in mothers remains unknown. In this study, in addition to an increase in serum serotonin levels, the serotonin and Tph1 levels in the maternal hearts also increased during pregnancy, suggesting a significant role for serotonin in maternal hearts.
Pregnancy induces electrocardiogram disturbances, such as a longer QT-interval, accompanied by downregulation of Ito,f and IK,slow.420 The expression of the cardiac Kv4.3 channel was down-regulated 3- to 5-fold, and was paralleled by a reduction in the transient Kv currents, a longer action potential, and prolongation of the QT interval. In this study, serotonin and Htr3 agonists shortened the QT interval and increased Kv currents in non-pregnant mice. The maternal heart significantly adapts to the circulatory needs of pregnancy. Interestingly Interestingly, the Htr3a antagonist ondansetron further decreased the Kv current in pregnant mice. While Kv4.3 downregulation leads to prolongation of action potential duration in cardiac hypertrophy and/or failure of rodents, it may not play a critical role in setting of cardiac action potential duration in human and canine. Therefore, it is still unclear whether the underlying mechanism and features of pregnancy-related QT prolongation in mice are similar to those in humans. In addition, Kv4.3 complexes with Kv channel interacting protein 2 (KChIP2), which is a Ca2+-binding EF-hand protein that regulates Kv4.3 inactivation gating.4 This raises the possibility that 5-HT3 receptor-mediated Ca2+ increase can modulate Kv4.3 currents via KChIP2. This finding suggests that serotonin compensates for QT prolongation during pregnancy by increasing Kv currents via Htr3. Consistently, our previous study showed that pregnant Htr3a−/− mice displayed a prolonged QT interval, compared to wild pregnant mice.10 As a study limitation, however, we used pre-pulse at −40 mV to inactivate voltage-gated Na+ currents. Kv4.3 is transient outward the K+ channel (Ito) whose inactivation kinetics is similar to voltage-gated Na+ channels. It is conceivable that Kv4.3 may be inactivated. In addition, 5-HT3 receptor is a Ca2+-permeable channel. Therefore, there is the possibility that it activates Ca2+-activated K+ currents. On the other hand, it is known that CaMKII interacts with Kv4.3 to regulate channel activity. These points argue that 5-HT3-mediated Ca2+ influx may have multiple effects on regulating action potential duration.
Serotonin increased Kv current densities via Htr3. However, although we demonstrated the interaction between Htr3a and Kv channels using co-immunoprecipitation, it remains unclear whether the activated Htr3 increase was directly or indirectly influenced Kv current densities. Also, we did not confirm whether increased Kv4.3 residence time at the plasma membrane is mediated via increased exocytosis or decreased endocytosis of the channel. Serotonin and Tph1 increased during pregnancy. Peripherally, serotonin is stored in platelets. However, we could not exactly identify the source of the serotonin that affected the heart.
Serotonin decreased QT intervals by increasing repolarizing currents, such as Kv current, via Htr3a in mouse hearts. During pregnancy, Htr3a-mediated Kv4.3 membrane trafficking was saturated. Elevated serotonin levels counterbalanced pregnancy-related QT prolongation by facilitating Htr3-mediated Kv currents. These results provide mechanistic insights into the hormonal control of ventricular repolarization during pregnancy in mice.


 XML Download
XML Download