

 PDF
PDF ePub
ePub Citation
Citation Print
Print



INTRODUCTION
Emerging infectious diseases (EIDs) are defined as newly appearing infections in a population or diseases that have previously existed, but are rapidly emerging in incidence or geographic range.12 EIDs are often initiated by pathogens with a change in pathogenicity, and these emerging pathogens are highly likely to be zoonotic, with a broad range of hosts or vector-borne.345 New human pathogens are especially likely to continue emerging, mainly from mammals and birds, for the foreseeable future. For these reasons, an effective global surveillance system for novel pathogens is needed. Due to the diverse nature of antigenic variation of emerging pathogens and their various strategies to evade the immune system, EIDs continue to pose difficult challenges to clinicians, public health professionals, and researchers globally.
An understanding of EID pathogenesis begins with the investigation of the immune response to emerging pathogens. An immune response is the consequence of an intricate and elaborate network of a large number of specialized cells and organs operating over space and time. In response to pathogens, an immune system must orchestrate the complex interactions and activities of a large number of diverse cell types, thereby recognizing and eliminating the pathogens. The ability to respond quickly and effectively to control invading pathogens is achieved by both the innate and adaptive immune systems. Innate immunity provides a fast and immediate sensing of pathogens, whereas adaptive immunity is responsible for a late and long-lasting protective immune memory that is able to respond quickly to future infections. The major goal of vaccines is to mimic these naturally induced memory responses by training the immune system with altered forms of a pathogen or components of pathogens that do not cause illness. Th-erefore, the success of vaccination largely depends on the quality of vaccine-induced immunogenicity.
Thus far, vaccines have been prepared by traditional methods, involving isolating infectious agents, attenuating or inactivating them, and exposing them to the immune system.6 Although this traditional approach has proven very efficient against pathogens with relatively low antigen variability, such as smallpox, polio, measles, mumps, and rubella, these approaches are not optimal to provide adequate protection against recently appearing EIDs, such as avian influenza virus infection, Ebola hemorrhagic fever, and dengue fever.78 Thus, novel systemic approaches are required to predict protective immune responses from high throughput sequencing and bioinformatics analysis and to identify potential antigenic targets to guide and accelerate vaccine development.
In recent years, systems biology has rapidly emerged as a new discipline.9 As researchers move from a traditional reductionist approach to a systemic approach, there are rapidly increasing amounts of novel omics data and other relevant data available. Recent advances in next-generation sequencing (NGS) and high-throughput technologies have enabled researchers to comprehensively assess whole genomes, transcriptomes, proteomes, and metabolomes of cells and tissues. These systemic approaches are being applied to the field of immunology and vaccine development, since the integration of genomics, transcriptomics, and metabolomics can give a global picture of nearly 30000 genes, proteins, and cells participating in the immune response to pathogens or vaccination.10111213
In this review, we highlight the major EIDs that have had a significant global impact and for which vaccines do not currently exist, and we summarize recent EIDs in South Korea. Moreover, we discuss various systems biology approaches and examine how these strategies offer significant potential for discovering new biomarkers of pathogenicity or predictors of protective immunity. Overall, we suggest that a systems biology approach would likely enhance our understanding of the pathogenesis of newly emerging pathogens and accelerate the development of next generation vaccines against EIDs.
EMERGING INFECTIOUS DISEASES
Recent global outbreaks of EIDs, such as Ebola or Zika virus (ZIKV) diseases, reminded the world of the imminent threat of unexpected pathogens. EIDs can result from infectious agents that have recently appeared or have existed, but are rapidly increasing in incidence.1415 Major pathogens include highly pathogenic avian influenza (HPAI) virus, severe fever with thrombocytopenia syndrome (SFTS) virus (SFTSV), ZIKV, dengue virus (DENV), and Middle East respiratory syndrome coronavirus (MERS-CoV).
Southeast Asia has been the epicenter of many emerging diseases in the past decade.16 Table 1 shows the characteristics of recently occurred or current EIDs occurring in Southeast Asia in the 21st century. Such diseases pose a major threat to public health and have the potential to cause a high incidence of fatality among the global community. Therefore, further investigation into the emergence and pathogenesis of EIDs is required for public health and safety.
There are several factors contributing to the emergence of new pathogens: microbial adaptation and genomic change, increased frequency of international travel and commerce, and climate change.17 Given that microbial adaptation is driven by the evolution of microbes to survive in diverse environments, microbes have evolved to change their genomes. RNA viruses carry RNA as their genetic material and can quickly adapt to and exploit different hosts because of the high error rates of the viral enzymes (RNA-dependent RNA polymerases) during their replication.18 The segmented nature and faulty replication mechanisms of Orthomyxoviruses promote genetic variability and frequent re-assortment events, resulting in genetically diverse and novel genomic constellations in infected hosts.19 As an example, hemagglutinin surface proteins of the influenza A virus undergo antigenic drift and shift, acquiring new infectivity and evading recognition by the immune system.20 Another important consequence of the unstable genomic make-up of RNA viruses is the potential for interspecies transmission between animal hosts and humans. Accordingly, the recent emergence of avian influenza viruses could be devastating in an immunologically naïve population and pose a serious threat to public health.21
Another major factor responsible for EIDs is an increased incidence of international travel and commerce.2223 The frequent use of air travel has enabled increased globalization, and international tourism has expanded 1.5 times in the last decade. Many travelers can be infected by and become carriers of EID pathogens without their knowledge. A viral incubation period can frequently last more than 2 weeks; thus, it is impossible to successfully monitor travelers with EIDs at an early stage of infection.24 Surveillance of travelers with respiratory illness is especially difficult, because respiratory viruses can propagate easily and cause outbreaks in non-endemic disease areas. For example, in 2015, South Korea experienced a MERS outbreak, which was initiated by a man who had traveled from Bahrain, which is located in the Middle East.25 Despite South Korea being a non-endemic area for MERS, 186 cases were reportedly infected with MERS-CoV during the 2015 outbreak, and this number was the highest among MERS non-endemic areas.26
Lastly, climate change can contribute to ecological changes and, thus, contribute to the emergence of EIDs. Over the last 10 years, the temperature of the earth has increased by approximately 0.4℃, and this global warming has contributed to climate change throughout the world. A warmer climate can provide a more favorable environment for the survival and completion of the vector life cycle.27 In particular, the mosquito population has been widely established across Europe, as well as tropical regions, such as Africa and South Asia.28 As an example, prior to 1970, only nine countries had experienced dengue fever, which is transmitted by mosquitoes and occurs mostly in tropical regions. However, today, cases of DENV-associated disease can be found in more than 125 countries.29 Moreover, climate change is frequently invoked as a primary cause of expansions in incidences of tick-borne diseases, particularly Lyme disease.30 A wide distribution of tick vectors increased the prevalence of tick-borne pathogens, given that rising temperatures accelerate the cycle of development, the production of eggs, and the density and distribution of the tick population.31 Therefore, it is necessary to explore how climate change will alter the distribution, dynamics, and risk of vector-borne diseases.
EIDS IN SOUTH KOREA
Highly pathogenic avian influenza infection
In South Korea, there are currently several major EIDs, such as HPAI infection, SFTSV infection, MERS-CoV infection, and DENV/ZIKV-associated diseases, that could pose great risks to public health in the near future. Since 2003, outbreaks of HPAI virus have been alarming, because they have caused significant economic loss and public health concerns.32 The HPAI virus can undergo rapid evolution by gene mutation, reassortment, and homologous recombination in avian species and vertebrate reservoir hosts.33 Although there is no evidence thus far to suggest a direct transmission of HPAI virus to humans in Korea, concerns remain due to the potential for avian influenza viruses circulating in poultry to become transmissible between species and to directly infect humans. In addition, due to the recent increase in zoonotic infections in poultry and persistent human infections in China, influenza A (H7N9) virus has remained a public health threat. Moreover, in February 2013, cases of human infection with a novel, lowly pathogenic H7N9 virus were reported in the Anhui and Shanghai regions of eastern China, and as of April 2017, the total number of H7N9 cases has exceeded 1344, with 511 deaths.3435
Severe fever with thrombocytopenia syndrome virus infection
SFTS infection is an emerging tick-borne hemorrhagic fever that was first described in rural areas of China. The causative agent, SFTSV, is a novel phlebovirus in the Bunyaviridae family. SFTS cases were first reported in China (2009), Japan (2012), and Korea (2013).36373839 The major clinical signs and symptoms of SFTS include high fever, gastrointestinal symptoms, thrombocytopenia, leukocytopenia, and multi-organ dysfunction, with an average case-fatality rate of 10–30%. Moreover, bone marrow suppression and hemophagocytic histiocytes are commonly found in Korean patients with SFTS infection.40 Despite no available treatment for SFTS infection yet, early plasma exchange implementation has proved to be beneficial.41 Moreover, SFTSV is believed to be transmitted through tick bites; however, there are a few suspected cases of human-to-human transmission in China and Korea.42434445 Although human-to-human transmission is considered very rare, it is urgently necessary to develop effective diagnostic and preventative tools to protect against SFTSV and maintain continuous surveillance, because the virus is thought to spread via blood or mucosal secretions.
Middle East respiratory syndrome coronavirus infection
MERS-CoV infection is a respiratory disease caused by MERS-CoV, a positive single stranded RNA virus.46 Although the detailed mode of transmission and period of virus shedding from infected patients are not fully understood, a nosocomial infection process is anticipated as the main transmission process.47 MERS-CoV was first identified in the Middle East where dromedary camels were found to carry MERS-CoV without any respiratory symptoms.48 The fatality rate of MERS-CoV infection in humans is approximately 20–50%, and its symptoms include fever, cough, and breathing difficulties.48 Since 2012, about 30 countries, including Saudi Arabia, China, Malaysia, and Jordan, have reported MERS outbreaks, with Saudi Arabia reporting approximately 80% of all outbreaks.49 Surprisingly, South Korea, which is a non-endemic area for MERS, experienced MERS outbreaks in 2015.26 It is thought that a traveler returning from Bahrain carrying the MERS virus caused an outbreak of 186 cases and 36 deaths, which had a substantial impact on the South Korean economy.26 Given the continuum of emergence and the large magnitude of MERS outbreaks in various regions, further studies are required to bolster capabilities for timely detection and effective control and prevention of MERS-CoV.
ZIKV/DENV-associated infection
As a consequence of global warming, ZIKV/DENV-associated diseases are potential EIDs that could arise in South Korea. DENV and ZIKV are mainly transmitted by Aedes albopictus and Aedes aegypti mosquitoes, respectively. Dengue fever is a major public health concern in many parts of the tropics and subtropics, and four different serotypes are currently co-circulating in most dengue-endemic areas.50 The pathogenesis of dengue fever is very unique in regards to antibody-dependent immune enhancement associated with DENV. Whereas primary infection usually confers durable, if not life-long, protection against re-infection by a homologous DENV serotype, secondary infection by viruses of a heterologous DENV serotype can frequently occur and result in severe pathology of the host.5152 Thus, the unique complexity of DENV pathogenesis and its effect on immune enhancement pose a great challenge to the development of a safe and effective DENV vaccine. Even though South Korea is not a region where dengue is epidemic, there have been many imported DENV infection cases in South Korea, and their number is increasing due to increased traveling to Southeast Asia.5354
Likewise, ZIKV is an another emerging mosquito-borne pathogen which belongs to the family Flaviviridae and is closely related to other pathogens of public health importance, including Yellow Fever virus, DENV, Japanese Encephalitis virus, and West Nile virus. The first case of human infection by ZIKV was reported in Uganda in 1952.55 Usually, ZIKV infection results in asymptomatic clinical features, but in some cases, ZIKV infection can lead to fatal microcephaly in infants and Guillain-Barre syndrome in adults.56 With increasing cases of ZIKV infection in America, many Asian countries, including India, the Philippines, Taiwan, and Vietnam, have reported ZIKV outbreaks, as of 2016.57 Although South Korea is considered a non-endemic region for Aedes aegypti-transmitted diseases, the first patient infected with ZIKV was reported in South Korea in 2016. The patient had stayed in Brazil for 3 weeks, and upon arrival in South Korea, complained of various symptoms, including fever, eyeball pain, and maculopapular rash.58 Given that Asia is endemic for ZIKV infection and the climate of South Korea is changing each year, it will be only be a matter of time before mosquito-borne EIDs become endemic in South Korea. Thus, we need to prepare for protection against mosquito-borne EIDs.
As summarized above, in addition to virus genetic variation, complex environmental factors, including ecological and behavioral influences, can be important contributing components of the recent emergence of viral diseases. Recent Ebola and Zika outbreaks have shown that the world is unprepared to detect local outbreaks and respond quickly enough to prevent them from becoming global pandemics. Given the great concern for public health owing to the speed and increased volume of global transportation, the ability to rapidly develop and deliver vaccines when new ‘unknown’ diseases emerge offers our best hope to outpace EID outbreaks, save lives, and avert disastrous economic consequences.
LEARNING FROM SYSTEMS BIOLOGY APPROACHES
As mentioned above, due to ecological, environmental, and demographic factors that increase exposure to emerging pathogens, EID cases are increasing in prevalence globally. Together with the ongoing evolution of viral and microbial variants and selection for drug resistance, these factors will contribute to the continued emergence of EIDs. Therefore, there is an urgent need for effective surveillance and control strategies, as well as rapid vaccine development. Early warning of EIDs depends on the ability to identify the unusual as early as possible. Information is, therefore, essential and this can be achieved using systems biology tools.
Systems biology is an approach to understanding living systems that focuses on modeling diverse types of high-dimensional interactions to develop a more comprehensive understanding of complex phenotypes manifested by the system.10 Systems biology tools enable us to develop and validate predictive models of infectious disease initiation, progression, and outcomes. These models can be derived from the study of the architecture and dynamics of systems-wide host/pathogen molecular interaction networks during infection. Ideally, integrated datasets generated from a combination of high-throughput multi-omics data (transcriptomics, proteomics, metabolomics, lipidomics, etc.) can be used to construct predictive models of the networks and dynamic interactions between the biological components of the complex pathogen-host system.59
In particular, through years of dedicated effort, high-throughput RNA sequencing (RNA-seq) technology, which is a powerful way to profile the transcriptome with great efficiency and high accuracy, has been employed in various viral infections and diseases.60616263646566 These studies proved that RNA-seq technology has the potential to reveal the dynamics of pathogen genome alteration and systemic changes in host and pathogen gene expression during the process of infection. Thus, if this approach is applied to EID research, information can help to accelerate uncovering the pathogenesis of novel infections and the interaction mechanism of emerging pathogens. As a notable example, several groups have taken advantage of RNA-seq technology to provide novel insights into how ZIKV infection may cause microcephaly in a short period of time. Nowakowki, et al.67 using single-cell RNA-seq and immunohistochemistry, first reported that AXL, a viral receptor for ZIKV, is highly expressed in human radial glial cells, astrocytes, endothelial cells, and microglia in the developing human cortex and in progenitor cells in the developing retina. Additionally, Onorati, et al.68 analyzed ZIKV-infected neuronal progenitor cells to identify transcriptional changes caused by the virus. Using techniques including single-cell RNA-seq, they described the derivation and characterization of neocortical and spinal cord neuroepithelial stem cells to model early human neurodevelopment and ZIKV-related neuropathogenesis. These data may lead to significant advances in efforts to develop a vaccine against ZIKV.
As discussed above, more extensive knowledge of the factors underlying EID pathogenesis can help with the development of more effective prevention strategies. If we are to protect ourselves against EIDs, the essential first step is to establish effective global disease surveillance to give an early warning of emerging infections. This can be achieved by rapid, portable, and inexpensive high-throughput screening of EID pathogens. Furthermore, systems-level informatics will not only reduce the time required for experimental immunology and vaccinology, but also promote identification and rational design of novel vaccine candidates and the determinants of vaccine-induced immunity. This will prompt the escalation of control measures where necessary and facilitate outbreak tracking.
SYSTEMS VACCINOLOGY APPROACHES
Vaccines are one of the greatest successes in the history of mankind. However, despite a global effort to develop vaccines against major infectious diseases, the correlates of protection are still poorly defined for many existing vaccines. Moreover, pathogens can evade and suppress immune responses, and human genetic variability can contribute to host susceptibility to infectious diseases or vaccine responsiveness. Although the eradication of smallpox is one of the many successes of global immunization programs, many vaccine challenges remain, especially for the design of effective vaccines for major killers, such as AIDS, dengue fever, and malaria.69 Furthermore, there is an urgent need to develop vaccines for EIDs, including the diseases mentioned above and neglected diseases, such as those caused by Chikungunya, Hendra, and Nipah viruses.
Recent studies have used systems biology approaches to identify molecular networks that orchestrate immunity to vaccinations in humans and potentially define correlates of protection10117071727374757677787980 The first attempt at this approach was made with one of the most successful vaccines ever developed, the yellow fever YF-17D vaccine.7081 As a live attenuated virus vaccine, a single shot of the yellow fever vaccine can confer protection in more than 90% of vaccinees, and it results in long-lasting immunity, a strong neutralizing antibody response, and a robust T cell response. Two independent groups have simultaneously published gene expression profiling data of blood samples from vaccinees within a few days after vaccination.7082 These molecular signatures helped researchers predict the magnitude of vaccine-induced immunity and provided novel insights into the dynamic nature of the innate and adaptive immune responses to YF-17D vaccine. In particular, Querec, et al.70 reported that EIF2AK4 (also known as GCN2), a known sensor of low amino acid levels inside the cell, was frequently found in the blood of vaccinees and was strongly correlated with the magnitude of the later CD8 T cell response to YF-17D vaccine.
Subsequently, systems biology approaches have been extended to influenza and other vaccine studies. Identifying molecular mechanisms of influenza vaccine-induced immunity was more challenging than the yellow fever vaccine study, given that the variation in antigenic match between the vaccine and circulating influenza strains can contribute to vaccine efficacy. Nakaya, et al.71 compared transcriptomic data for those given either the trivalent influenza vaccine (TIV) or the live attenuated influenza vaccine (LAIV). Both vaccines induced the expression of genes involved in innate immune pathways, although LAIV induced higher expression of several interferon-related genes, similar to another live attenuated vaccine, YF17D. Therefore, these data demonstrated that live and dead influenza vaccines clearly result in differential vaccine-induced immunity.
Among many factors that determine the efficacy of vaccines, the age of vaccinees is the most important. In fact, older adults and children are most vulnerable to severe and often lethal influenza virus infection, due to decreased vaccine efficacy. Thus, the use of adjuvants can be helpful to improve the efficacy of influenza virus vaccines for children and older adults.838485 In support of this idea, Nakaya, et al.75 conducted blood transcriptome analysis in children less than 2 years old to compare the immunogenicity of TIV with and without MF59 adjuvant, an oil-in-water adjuvant known to increase the magnitude of the antibody response to inactivated vaccines. Interestingly, the more potent antibody responses generated in subjects receiving MF59-adjuvanted vaccines correlated with strong transcriptional signatures of interferon signaling network in blood samples.
In addition to transcriptome analysis, other systems-level approaches, such as proteomics, metabolomics, and lipidomics, can provide greater depth in evaluating correlates of vaccine-induced immunity. The latest research by Dr. Pulendran's group analyzed vaccine-induced immunity in multi-scale ways in different age groups of healthy adults immunized with shingles vaccine, Zostavax.77 The authors presented an integration data of peripheral blood mononuclear cells (PBMC) transcriptomics, flow cytometry of blood cell populations, plasma cytokine analysis, and metabolomics. The integration of these data revealed transcriptomic association with metabolic correlates of vaccine immunity. Interestingly, this analysis revealed sterol regulatory binding protein-1 and its targets as potentially key integrators of antibody and follicular helper T cell responses. Therefore, these data suggest that blood cells potentially regulate gene transcription in response to metabolic cues, suggesting the value in integration of diverse systems-level data.
Given that a vaccine usually initiates a broad, polyfunctional, and persistent immune response integrating all effector cells of the immune system, it is important for scientists to understand the complexity and dynamic and diverse quality of the immune response.86 Therefore, innovative tools, such as using single cell technologies to ‘omics’ approaches, have enabled researchers to quantitatively examine entire classes of molecules at a single cell level, and should further facilitate our understanding of the functional diversity of the immune response. As shown in Fig. 1, using systems biology tools for increased understanding of vaccine-mediated immunity can help scientists be better positioned to identify correlates of vaccine immunogenicity and make progress towards developing new and enhanced vaccines and therapeutics.
CURRENT LIMITATIONS AND CHALLENGES
Thus far, we have reviewed recent advances in the fields of systems immunology and vaccinology, and presented how systems biology-based platforms can be applied to accelerate a deeper understanding of molecular mechanisms of immunity against EIDs. Despite recent advances, there remain computational and biological challenges regarding systems biology approaches. Because the immune response to a vaccine can be affected by a wide range of confounding factors, including cellular, population, molecular, individual, and technical factors, scientists face many challenges and potential limitations on the analysis of systems-level informatics. These challenges and limitations are described in Fig. 2.
Before conducting a large-scale systems biology approach, one should consider the following limitations. Although previous studies involving mathematical modeling of the dynamics of immune responses can provide clues for optimal experimental design, systems-level experiments should be very carefully designed to control for all variables in terms of time points. Furthermore, given that an immune response occurs as a consequence of coordinated events among many immune cells over space and time, it is important to understand how immune cells respond at both the cell population and the single cell level. Single-cell sequencing allows researchers to examine the genomes or transcriptomes of individual cells, thus providing a high-resolution view of cell-to-cell variation. This technique can be used to assess the individual contribution of single cells in complex tissues by profiling the transcriptome in an unbiased manner. In addition, technological problems during systems biology approaches can be resolved by the solutions shown in Table 2.
Additionally, despite the high costs of high throughput analysis and limited numbers of human samples, to minimize confounding factors, it is best to maximize sample sizes to achieve appropriate statistical power. Besides sample size, another major confounding variable from a systems vaccinology approach is that the observed transcriptomic changes may result from new induction of gene expression or may simply reflect the changing cellular composition of PBMC compartments. Given that it is impossible to obtain cellular or tissue samples from lymph nodes or spleens of healthy human subjects post vaccination, vaccine efficacy tests must be performed on PBMCs drawn pre- and post-vaccination. This limits the optimal understanding of vaccine-induced protective immune responses. One should keep in mind that PBMCs include mixed and dynamic cell populations, and thus, there may be dynamic events occurring within lymphoid compartments of PBMCs.8788 Therefore, it will be more informative and precise to determine the transcriptome in sorted cell subsets, such as B cells, T cells, natural killer cells, macrophages, and dendritic cells.
REVERSE VACCINOLOGY-BASED VACCINE DESIGN
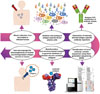
Emerging technologies, such as structure-based antigen design and synthetic vaccinology, have heralded in a new era of vaccine discovery and development. As an example, a reverse vaccinology process was recently applied, based on the notion that protective antigens could be identified by determining the antibody repertoire from infected or vaccinated individuals.8990 Fig. 3 demonstrates the identification of novel and the most effective epitopes using NGS and high-throughput robotic crystallization platforms to enable more precise prediction of epitope targets for efficient antibody generation. Web-based epitope databases, vaccine design resources, and protein structure comparison resources can help to improve our understanding of immune responses and the escape and evolution of pathogens.91 As an example, there have been massive antibody engineering efforts to improve the breadth and potency of anti-HIV-1 antibodies.92 Given the importance of neutralizing antibodies for vaccine protection, the dissection of antibody variable regions using reverse vaccinology tools is a rational approach to design vaccines and will ultimately lead to more effective vaccines for EIDs.
SUMMARY AND FUTURE DIRECTIONS
Recent advances in systems biology-based technologies have enabled scientists to investigate the immune responses induced by pathogens and vaccines at a greater depth than ever before. More importantly, these advances are facilitating the identification of robust molecular and cellular signatures of protective immunity, which can help to generate diagnostic tools that reduce the length and cost of current clinical trials. When these systems approaches are applied to EIDs, they will enable more rapid and rational design of vaccine development. Thus, in this developing field, vaccinologists, immunologists, bioinformaticians, and systems biologists must work hand-in-hand to advance our understanding of the molecular mechanisms by which vaccines induce protective immunity and help drive development of the next generation of vaccines to prevent EIDs.




 XML Download
XML Download