

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Schizophrenia is a devastating mental disorder and is characterized by several symptoms related to abnormalities in perception and expression, such as delusion, hallucination, and disorganized thinking.1 Although evidence on gender differences in schizophrenia development is inconclusive,2 the prevalence of the disease is different between the sexes, with about a 1.4-fold higher risk in men than in women.3 The heritability of schizophrenia is reported to be around 80%, suggesting that genetic factors are strongly involved in the risk for the disorder.4 The putative susceptible genes for schizophrenia are dystrobrevin-binding protein 1, neuregulin 1 (NRG1), D-amino-acid oxidase (DAOA), and regulator of G-protein signaling 4 (RGS4).5 Recently, chromogranin B (CHGB) has also been identified as a candidate gene influencing the risk of schizophrenia,678910 although robust evidence has not been reported. In a genome-wide association study (GWAS) performed in a Japanese population, a genetic variant (microsatellite: D20S95) near CHGB was discovered as a potential genetic marker for schizophrenia development.11 In addition, our GWAS of schizophrenia in a Korean population (in submission) has identified three markers of CHGB that were significantly associated with schizophrenia (minimum p=3.2×10-6 at rs2821). Inspired by these results, this study performed a replication study of the association between schizophrenia development and CHGB using additional single nucleotide polymorphisms (SNPs).
Accumulating evidence indicates that synaptic disturbance or damage may be important in the etiology of schizophrenia. Granins (secretogranins/chromogranins) are a family of soluble proteins that are stored in and released from the secretory large dense-core vesicles of the synapse. The CHGB, also known as secretogranin I, is an acidic glycoprotein that is present in secretory granules of neuroendocrine cells and neurons.12 In genetic association studies performed in Asian populations (Japanese and Chinese) genetic makers in CHGB (including several non-synonymous variants of rs6133278, rs910122, rs236152, rs236153, and rs74621755) have been reported to be associated with the risk of schizophrenia.91013
In order to investigate potential genetic markers of CHGB for schizophrenia, we analyzed the associations of polymorphisms in CHGB with the risk of the disease in a Korean population.
MATERIALS AND METHODS
Study subjects
A total of 310 patients with schizophrenia (mean age=44.7 years; range=23–73 years; 185 males and 125 females) were recruited from Seoul National University, Jinju Mental, Soonyoung University, Hadong Wooridle, and Keyo Hospitals. A total of 604 healthy controls (mean age=48.79 years; range=8–84 years; 254 males and 350 females) were simultaneously recruited from an unselected population who had come in for routine health checkups in the same regional areas. Healthy controls were recruited from Seoul National University and Hallym University Hospitals. Trained psychiatrists diagnosed schizophrenia patients based on the criteria set forth by the Diagnostic and Statistical Manual of Mental Disorders, fourth edition (DSM-IV).14 Patients with complicating diagnoses of mental retardation, organic brain damage, neurological disorders, drug or alcohol abuse, autoimmune disorders, and low comprehension skills were excluded from the study. To ensure no history and present evidence of psychiatric illness, each control subject underwent additional evaluation by trained psychiatrists using the Structured Clinical Interview from DSM-IV, non-patient edition. The ethnicity of all patients and healthy controls was Korean. Informed written consent was obtained from all subjects before blood was drawn. The study protocol was approved by the Institutional Review Board of each hospital.
SNP selection and genotyping
Candidate SNPs of CHGB were selected from genotype data from Japanese and Han Chinese populations in the 1000 Genomes database (http://browser.1000genomes.org/index.html) based on the following conditions: 1) minor allele frequency (MAF) >5%, 2) linkage disequilibrium (LD) status [LD coefficient (r2) >0.98], 3) position within the gene, and 4) amino acid changes. In addition, previously reported SNPs were included. A total of 15 SNPs of CHGB (rs226137, rs236139, rs236141, rs16991480, rs76791154, rs236145, rs446659, rs6085323, rs6085324, rs6133278, rs910122, rs881118, rs742711, rs74621755, rs2821) were genotyped in 310 schizophrenia patients and 604 healthy controls using the TaqMan assay on the ABI Prism 7900HT (Applied Biosystems, Foster City, CA, USA). Genotyping quality control was performed in 10% of the samples by duplicate checking (rate of concordance in duplicates >99.5%). Selected SNPs and their probe information are available in Supplementary Table 1 (only online).
Statistical analysis
LD was obtained using the Haploview v4.2 software from the Broad Institute (http://www.broadinstitute.org/mpg/haploview), with examination of Lewontin's D′ (|D′|) and the r2 between all pairs of bi-allelic loci.15 Haplotypes (hts) were estimated using the PHASE software.16 Comparison of genotype distributions between schizophrenia patients and healthy controls was carried out with a logistic regression model adjusted for age (continuous value) and sex (male=0, female=1) as covariates using SAS, version 9.4 (SAS Inc., Cary, NC, USA). The effective number of independent marker loci was calculated for multiple testing corrections using SNPSpD (https://gump.qimr.edu.au/general/daleN/SNPSpD/), a program that is based on the spectral decomposition of matrices of pair-wise LD between SNPs.17 The total sum of the tested independent marker loci in the gene was calculated as 10.9139, which was applied to correct for multiple testing (p value×10.9139).
RESULTS
Genotyping and haplotypes of CHGB polymorphisms
A total of 15 CHGB genetic variants, including six non-synonymous SNPs (rs6085324, rs6133278, rs910122, rs881118, rs742711, and rs74621755), were selected from Asian populations (Chinese and Japanese) in the 1000 Genomes database, based on the selection criteria, and were genotyped (Fig. 1A). Detailed information about 15 investigated polymorphisms in this study (such as allele, position, MAF, heterozygosity, and Hardy-Weinberg equilibrium) is presented in Table 1. LDs among SNPs were measured by calculating |D′| and r2 values. Among the investigated genetic variants, one SNP (rs236137) was excluded from the LD block construction due to its low frequency (MAF <5%), whereas the other SNPs with MAF over 5% were used for LD block construction (Fig. 1B). The LD block was composed of five major hts with a frequency over 5% (Fig. 1C). Since the haplotype ht5 was equivalent to rs74621755, ht5 was excluded from further statistical analysis.
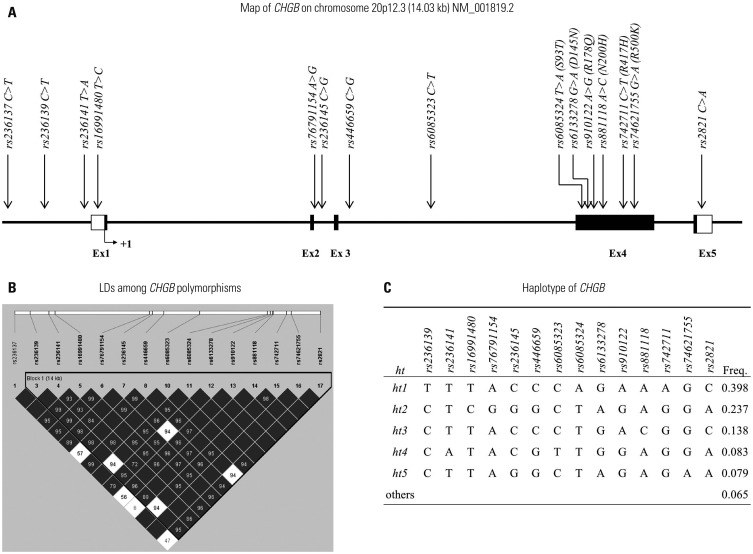
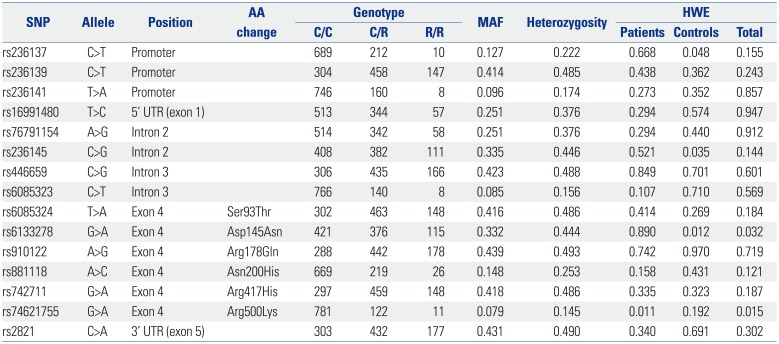
Fig. 1
Schematic physical map of CHGB. (A) Polymorphisms of CHGB investigated in this study. Black blocks indicate coding exons; white blocks, 5′- and 3′-untranslated regions. First base of translation site is denoted as nucleotide +1. (B) LDs among CHGB polymorphisms. (C) Haplotypes (hts) of CHGB in a Korean population. CHGB, chromogranin B; LD, linkage disequilibrium.
![]()
Table 1
Information of CHGB Polymorphisms among Subjects Investigated in This Study (n=914)
![]()
Associations of CHGB SNPs and haplotypes with schizophrenia
In order to investigate associations between CHGB genetic variants and schizophrenia in a Korean population, logistic regression analysis was performed. As a result, four SNPs, rs446659, rs6133278 (Asp145Asn), rs910122 (Arg178Gln), and rs2821, showed statistically significant associations with the risk of schizophrenia under the co-dominant model [minimum p=0.002 and odds ratio (OR)=0.72 at rs2821] (Table 2). Although significance at rs446659 and rs6133278 disappeared, significant signals at rs910122 and rs2821 were retained even after correction for multiple testing (pcorr<0.05). Due to different prevalences between men and women in schizophrenia, additional statistical analyses in both sexes were performed to determine the effect of sex on disease susceptibility. Interestingly, different significances in genetic associations were observed according to sex: five polymorphisms (rs236141, rs446659, rs6085323, rs910122, and rs2821) and one haplotype (ht4) showed significant associations with schizophrenia in male subjects (minimum p=0.002 and OR=0.64 at rs2821) (Table 2), even after the correction for multiple testing (minimum pcorr=0.02 at rs2821). However, no significant association was observed in female subjects (p>0.05).
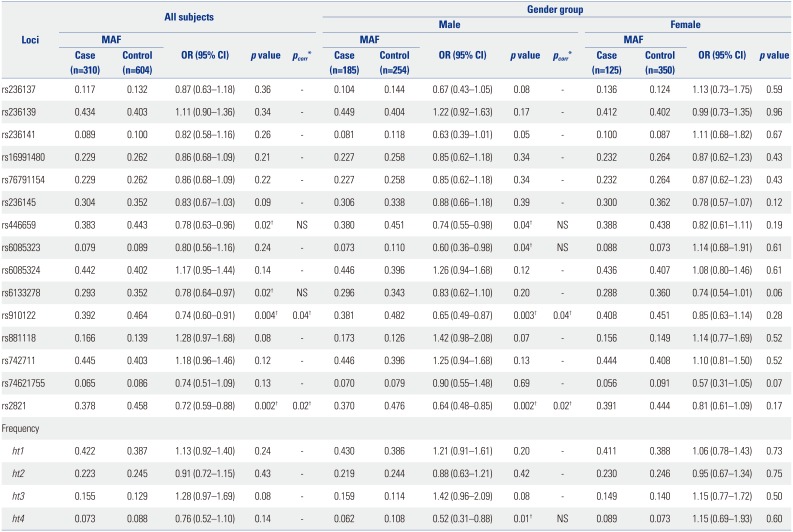
Table 2
Associations of CHGB Polymorphisms with Schizophrenia
MAF, minor allele frequency; OR, odds ratio; CI, confidence interval.
p value and OR are analyzed under the co-dominant model. Logistic analysis is adjusted by age and sex as covariates in case-control analysis and by age in gender group analysis. Haplotype 5 (ht5) is equivalent to rs74621755.
*pcorr value after multiple testing corrections with the effective number (10.9139) of independent marker loci, †Statistical significance of p<0.05.
![]()
In silico analysis
In order to predict the functional impact of the significantly associated polymorphisms (rs446659, rs6085323, rs6133278, rs910122, and rs2821; p<0.05) (Table 2), in silico analyses were performed. Among the investigated polymorphisms, several functional motifs were predicted on/nearby two non-synonymous SNPs (rs6133278 and rs910122) (Table 3) using the ELM resource for functional site prediction program (http://elm.eu.org/index.html). In particular, a motif (MOD_PIKK_1), including rs6133278 (D145N) is known as a phosphorylation site for phosphatidylinositol 3-kinase-related kinase (PIKK) family members that are recently implicated in brain development and functioning.
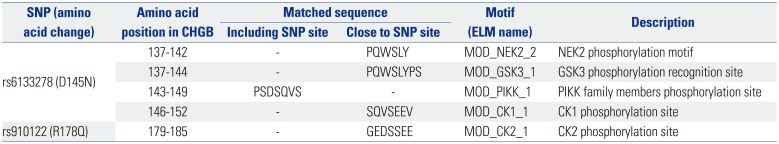
Table 3
Motif Search Analysis of rs6133278 and rs910122
![]()
DISCUSSION
Schizophrenia is a severe mental disorder that affects one's behaviors, emotions, and thinking. In addition, schizophrenia is considered to be a complex disorder affected by genetic, neurobiological, and environmental factors. Concentration of neurotransmitters (in particular, high dopamine, and low glutamate levels) has been considered as an important link between schizophrenia and altered brain function in terms of neurobiological process.1819 In addition, several studies have provided evidence that CHGB, a presynaptic protein that influences neurotransmitter release in neuroendocrine system, is differentially regulated in several brain neurological disorders, including schizophrenia.20212223 Recently, genetic associations on several neurotransmission related genes (Dysbindin, NRG1, DAOA, COMT, DISC1, and CHGB) with schizophrenia have been highlighted.2425 However, the exact causes of schizophrenia are not yet fully understood.
Among the proteins that participate in the neurotransmitter release process, CHGB has been reported to be associated with schizophrenia. The CHGB protein, which is also known as secretogranin I, is a widespread constituent of large dense-core vesicles and induces dopamine release by affecting the exocytosis of secretory pathway of hormones and/or neuropeptides. 26 The correlation between chromogranin and dopamine in schizophrenia has been emphasized in recent findings. A recent in vivo neuroimaging study has suggested that schizophrenia patients have increased synaptic dopamine concentrations in the brain when compared with a control group.27 Although an in vitro study reported that dopamine release might be increased by CHGB in a dose-dependent manner,28 another study also revealed that levels of CHGB were lower in the cerebrospinal fluid from schizophrenia patients than from healthy controls.20 Considering these previous reports and our results, the genetic effects induced by CHGB variants on schizophrenia in this study (minimum pcorr=0.02, OR=0.72 at rs2821) might have the possibility of being related to dopamine levels by influencing neurotransmission pathway.
Post-translational processes (such as phosphorylation, methylation, and glycosylation) that regulate protein function can be influenced by genetic variants, inducing an amino acid change. 2930 According to a previous report that examined changes in phosphorylation status induced by SNPs at a genome-wide level, the non-synonymous CHGB variant of rs910122 (Arg-178Gln) was reported to be a phospho-variant affecting adjacent CHGB phosphorylation sites by altering kinase activity.31 According to the study, the amino acid change from arginine to glutamine at CHGB rs910122 was shown to inhibit v-akt murine thymoma viral oncogene homolog kinase binding on nearby a serine residue at amino acid position 183 of CHGB. This result suggests that the non-synonymous variation may have a cis-acting effect on subsequent amino acid phosphorylation of CHGB, resulting in a considerable susceptibility to schizophrenia (OR=0.65 at rs910122). In addition, other protein kinases of PIKKs that were predicted to phosphorylate the motif including rs6133278 (Asp145Asn) in our in silico analysis (Table 3) have been reported to be associated with several neural activities, such as neuronal development and synaptic plasticity.3233 Among the PIKKs, the mammalian target of rapamycin kinase signaling is related to several cognitive disorders, including schizophrenia.34 Considering evidence that shows different phosphorylation patterns between schizophrenia patients and healthy controls,35 regulation of CHGB phosphorylation sites may play a role in schizophrenia development.
Previously, several studies have reported gender-specific differences in prevalence and development of schizophrenia.336 Moreover, gender-specific genetic markers in schizophrenia (such as rs2812393 and rs821616 of DISC1; rs175174 of ZDHHC8, for female patients) have been examined in recent genetic association studies.3738 Including the previously reported phospho-variant rs910122,31 different genetic effects of CHGB variants on the risk of schizophrenia were also observed depending on sex in this study (Table 2). Furthermore, considering that CHGB rs236141 with a nominal association with schizophrenia (p=0.05 and OR=0.63 in male subjects) has been found to be a gender-specific marker for blood pressure disease, 39 our findings suggest that CHGB could be a gender-specific marker for schizophrenia development.
In other studies on Chinese and Japanese populations for the association between CHGB SNPs and schizophrenia susceptibility, 913 rs910122 was observed as a consistent association signal with schizophrenia susceptibility in a Chinese population with significance, but not in a Japanese population. The rs2821 also showed significance in Chinese and Korean schizophrenia patients, but not in a Japanese population (Supplementary Table 2, only online). Instead, other CHGB SNPs were found to have significant signals in the Japanese patients, suggesting variations in CHGB may have different genetic effects depending on ethnicity.
There are study limitations of an insufficient number of samples and different proportions in age range (23–73 vs. 8–84) and sex ratio (M/F=185/125 vs. 254/350) between patients and controls, respectively, as well as no functional evaluation. Although logistic analysis of this study was adjusted by sex and age as covariates in case-control analysis and by age in gender group analysis, we also do not rule out a possibility that the significant association signals may have resulted from the study limitations. However, considering the different incidences of schizophrenia according to sex and age (such as male: female ratio of ~1.4:1 and earlier mean age at onset for schizophrenia in males),240 further replication studies in a larger cohort may be required.
In summary, this study replicated the association between CHGB polymorphisms and schizophrenia susceptibility. To our knowledge, this is the first study to identify the potential sex-specific association of CHGB genetic variants with schizophrenia. Despite study limitations, our findings may provide useful information on the genetic etiology of schizophrenia.
 XML Download
XML Download