

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Hypoxia, a condition of inadequate oxygen supply to cells and tissues, is frequently observed in almost all types of solid tumors, because of non-functional vasculatures and rapidly proliferating cancer cells outgrowing the existing supply. The hypoxic microenvironments inside tumors limit the effectiveness of radiotherapy and many cytotoxic drugs.1 In particular, cancer cells take advantage of their ability to adapt hypoxia to initiate a special transcriptional program that renders them more aggressive biological behaviors and poor clinical prognosis.2 The discovery of hypoxia-inducible factors (HIFs) as master driving forces of the cellular adaption to hypoxia has provided a fundamental molecular link to the dilemma.3 HIFs regulate a vast array of genes encoding proteins involved in cancer progression and treatment resistance.4 Therefore, HIFs have become targets for developing novel cancer therapeutics since early 1990s.5 A number of HIF inhibitors have been developed, and some of them are under investigation in clinical trials. This review summarizes the updated information in tumor HIF pathways, particularly the development of HIF inhibitors as potential anti-cancer agents.
Go to : 
TUMOR HYPOXIC MICROENVIRONMENTS
An adequate oxygen supply is crucial for cells of aerobic organisms to function and survive. The normal oxygen partial pressure in arterial blood is ~100 mm Hg (~13%) in a healthy body.6 In general, 8–10 mm Hg (~1%) is regarded as a critical oxygen partial pressure level that is associated with adverse effects of normal tissues caused by reduced oxygen consumption.7
In solid tumors, oxygen delivery to neoplastic and stromal cells is often reduced or even abolished due to severe structural abnormalities of microvessels and disturbed microcirculation. As a result, solid tumors frequently contain areas with very low oxygen pressure, occurring either acutely or chronically. These hypoxic regions are heterogeneously distributed within tumor masses and may even be located adjacent to vessels.8
Go to :
HYPOXIA-INDUCIBLE FACTORS AND THEIR STRUCTURE
Each HIF composes of an oxygen-sensitive α-subunit and a constitutively expressed β-subunit (also known as, aryl hydrocarbon receptor nuclear translocator).9 Up to now, three isoforms of HIFα have been identified, namely HIF-1α, HIF-2α, and HIF-3α.101112 This review focuses on HIF-1α and HIF-2α, since less is known about HIF-3α and no specific inhibitors targeting HIF-3 have been developed.13
HIF-1α and HIF-2α each has two transactivation domains (TAD) located at the NH2-terminal (N-TAD) and COOH-terminal (C-TAD). C-TAD interacts with p300/CREB-binding protein (CBP) co-activators to modulate gene transcription under hypoxia, while N-TAD is responsible for stabilizing HIFα against degradation.14 HIFα has an oxygen-dependent degradation domain (ODDD) overlapping N-TAD in their structures. The ODDD is important in mediating oxygen-regulated stability.15 Either HIF-1α or HIF-2α is degradable in an oxygen-dependent manner through the von Hippel-Lindau protein (pVHL) pathway, and is able to complex with HIF-1β to form a heterodimer (Fig. 1).16 HIFs bind to hypoxia-response elements (HREs) in the promoters of targeted genes (Fig. 1).12 Although HIF-1α and HIF-2α share a high degree of sequence identity (48% sequence similarity), a similar protein structure, and several common targets, they mediate unique patterns of gene regulation.1217 HIF-1α is ubiquitously expressed, while HIF-2α is by only certain cell-types and tumor-types. HIF-1α plays a dominant role in the response to acute hypoxia, whereas HIF-2α drives the response to chronic hypoxia, and the regulatory feedback of HIF-1α may be responsible for the selectivity.18
 | Fig. 1The HIF pathways and potential interfering points. Interfering points: a, HIFα mRNA; b, protein synthesis; c, protein stability and degradation; d, dimerization; e, DNA binding; f, transcriptional activity; g, Nuclear translocation. ARNT, aryl hydrocarbon receptor nuclear translocator (also called HIF-1α); GFs, growth factors; HIF, hypoxia-inducible factor; HREs, hypoxia-response elements; Hsp90, heat shock protein 90; PHD, prolyl hydroxylase; Ub, Ubiquitin; pVHL, von Hippel-Lindau tumor protein; HDAC, histone deacetylase; CBP, CREB-binding protein.
|
Go to :
MOLECULAR BASIS OF HIF REGULATION
HIFα is constitutively transcribed, translated and synthesized, and its stability and transcriptional activity are regulated by many growth factors and upstream cellular signaling pathways in oxygen-dependent and -independent manners.19 For instance, the expression of HIFα is mediated by phosphoinositide 3-kinase (PI3K) and mitogen-activated protein kinase (MAPK) pathways, which regulate phosphorylation of 4E-bindng protein-1 (4EBP1) and ribosomal S6 kinase (S6K), two essential factors for HIFα protein translation.20 The interaction of chaperone heat shock protein 90 (Hsp90) with HIFα is required for protein stability, particularly in the nascent process.21
Under normoxic conditions, HIFα is rapidly hydroxylated by prolyl hydroxylase (PHD), which mediates the binding by VHL ubiquitin ligase complex. The addition of poly-ubiquitin tags HIFα for the proteasomal degradation (Fig. 1). HIF-1α has a very short half-life (5 min) under normoxia.19 Under hypoxia, PHD loses its activity, thus preventing VHL binding and leading to the stabilization and accumulation of HIFα protein. HIFα translocates into the nucleus, where it heterodimerizes with HIF-1β to form HIF, which recruits the p300/CBP co-activators to form an active HIF transcription complex. This complex binds to the HREs on target genes and activates the transcription (Fig. 1). HREs are cis-acting elements, comprising the necessary but not sufficient HIF-binding sites with the consensus sequence RCGTG (R is purine A or G) and a HIF-ancillary sequence.1
Go to :
HIF-REGULATED GENES AND THEIR ROLES IN CANCER
Activation of HIF transcription leads to the upregulation of more than a hundred genes, which encode proteins participating in tumor angiogenesis, cell proliferation, survival, metabolism, invasion, and metastasis,1922 cancer stem cells,2324 therapy resistance,252627 and inflammation and immunity (Fig. 2).28 The regulation of target genes in hypoxic cells is mainly controlled by HIF-1 and HIF-2.12 Two of the well-studied functions of HIFs are regulating angiogenesis and glucose metabolism. Vascular endothelial growth factor (VEGF) is a powerful and major driver of tumor angiogenesis,16 and is commonly regulated by HIF-1 and HIF-2.12 HIFs also regulate genes involved in the glycolytic cascade, such as glucose transporters and lactate dehydrogenases.29 In addition, HIFs regulate other genes participating in decreasing the effectiveness of anticancer therapies such as radiation, chemotherapy and immunotherapy (Fig. 2).232425262728
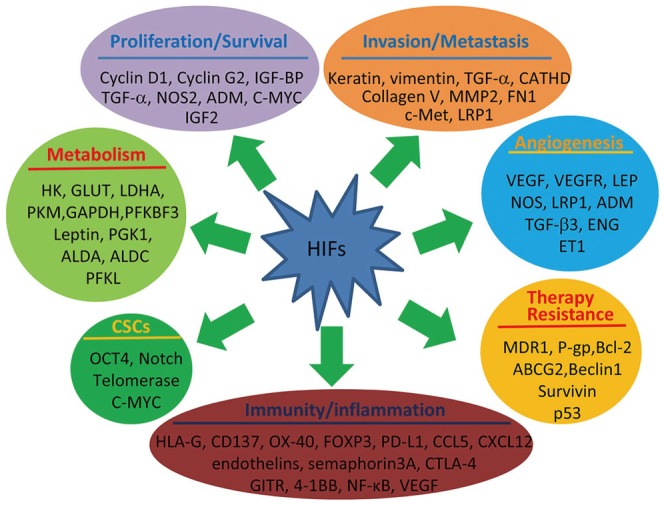 | Fig. 2Representative HIF-targeted genes and their roles in cancer progression and therapy resistance. ALDA, aldolase A; ALDC, aldolase C; ADM, adrenomedullin; ABCG2, ATP-binding cassette sub-family G member 2; CATHD, cathepsin D; CCL, chemokine (C-C motif) ligand; CTLA-4, cytotoxic T-lymphocyte-associated protein 4; CXCL, chemokine (C-X-C motif) ligand; ENG, endoglin; ET1, endothelin-1; FN1, fibronectin 1; FOXP3, forkhead box P3; GAPDH, glyceraldehyde-3-P-dehydrogenase; GITR, glucocorticoid-induced TNFR-related protein; GLUT, glucose transporter; HK, hexokinase; HIF, hypoxia-inducible factor; HLA-G, human leukocyte antigen G; IGF2, insulin-like growth factor 2; IGF-BP, insulin-like growth factor binding protein; LDHA, lactate dehydrogenase A; LRP1, LDL-receptor-related protein 1; MDR1, multidrug resistance 1; MMP2, matrix metal-loproteinase 2; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; NOS2, nitric oxide synthase 2; OCT4, octamer-binding transcription factor 4; PD-L1, programmed death-ligand 1; PFKBF3, 6-phosphofructo-2-kinase/fructose-2,6-biphosphatase-3; PFKL, 6-phosphofructokinase, liver type; PGK1, phosphoglycerate kinase 1; PKM, pyruvate kinase M; TGF, transforming growth factor; VEGF, vascular endothelial growth factor; VEGFR, VEGF receptor.
|
Go to :
HIF INHIBITORS FOR CANCER THERAPY
A number of HIF inhibitors have been synthesized or discovered for treating cancer, particularly for advanced and refractory solid tumors. They inhibit the expression and/or functions of HIF-1α, or HIF-2α, or both, through direct and indirect mechanisms. Those inhibitors which target the upstream pathways (such as the PI3K/AKT/mTOR pathways) or the downstream pathways (such as anti-VEGF-therapy) are not discussed in this review, since they are known for their other activities. Based on the putative mechanisms of action, HIF inhibitors can be tentatively classified into six types as below; agents modulate the expression, protein synthesis, protein accumulation and degradation, dimerization, DNA binding and transcriptional activity of HIFs, respectively (Fig. 1). However, the classification may not be accurate, because some agents target more than one mechanism and it may be inappropriate to include them in a definite category. The targets and mechanisms of HIF inhibitors (Table 1) and some evaluated in clinical trials [Data were obtained from National Clinical Trial (https://clinicaltrials.gov)] are summarized.3031323334353637383940414243444546474849505152535455
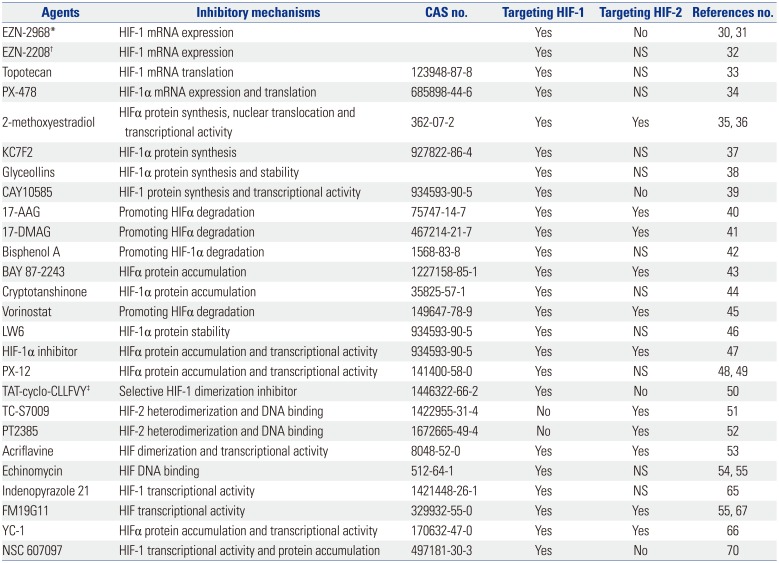
Table 1
The Targets and Mechanisms of HIF Inhibitors for Cancer Therapy
| Agents | Inhibitory mechanisms | CAS no. | Targeting HIF-1 | Targeting HIF-2 | References no. |
|---|---|---|---|---|---|
| EZN-2968* | HIF-1 mRNA expression | Yes | No | 30, 31 | |
| EZN-2208† | HIF-1 mRNA expression | Yes | NS | 32 | |
| Topotecan | HIF-1 mRNA translation | 123948-87-8 | Yes | NS | 33 |
| PX-478 | HIF-1α mRNA expression and translation | 685898-44-6 | Yes | NS | 34 |
| 2-methoxyestradiol | HIFα protein synthesis, nuclear translocation and transcriptional activity | 362-07-2 | Yes | Yes | 35, 36 |
| KC7F2 | HIF-1α protein synthesis | 927822-86-4 | Yes | NS | 37 |
| Glyceollins | HIF-1α protein synthesis and stability | Yes | NS | 38 | |
| CAY10585 | HIF-1 protein synthesis and transcriptional activity | 934593-90-5 | Yes | No | 39 |
| 17-AAG | Promoting HIFα degradation | 75747-14-7 | Yes | Yes | 40 |
| 17-DMAG | Promoting HIFα degradation | 467214-21-7 | Yes | Yes | 41 |
| Bisphenol A | Promoting HIF-1α degradation | 1568-83-8 | Yes | NS | 42 |
| BAY 87-2243 | HIFα protein accumulation | 1227158-85-1 | Yes | Yes | 43 |
| Cryptotanshinone | HIF-1α protein accumulation | 35825-57-1 | Yes | NS | 44 |
| Vorinostat | Promoting HIFα degradation | 149647-78-9 | Yes | Yes | 45 |
| LW6 | HIF-1α protein stability | 934593-90-5 | Yes | NS | 46 |
| HIF-1α inhibitor | HIFα protein accumulation and transcriptional activity | 934593-90-5 | Yes | Yes | 47 |
| PX-12 | HIFα protein accumulation and transcriptional activity | 141400-58-0 | Yes | NS | 48, 49 |
| TAT-cyclo-CLLFVY‡ | Selective HIF-1 dimerization inhibitor | 1446322-66-2 | Yes | No | 50 |
| TC-S7009 | HIF-2 heterodimerization and DNA binding | 1422955-31-4 | No | Yes | 51 |
| PT2385 | HIF-2 heterodimerization and DNA binding | 1672665-49-4 | No | Yes | 52 |
| Acriflavine | HIF dimerization and transcriptional activity | 8048-52-0 | Yes | Yes | 53 |
| Echinomycin | HIF DNA binding | 512-64-1 | Yes | NS | 54, 55 |
| Indenopyrazole 21 | HIF-1 transcriptional activity | 1421448-26-1 | Yes | NS | 65 |
| FM19G11 | HIF transcriptional activity | 329932-55-0 | Yes | Yes | 55, 67 |
| YC-1 | HIFα protein accumulation and transcriptional activity | 170632-47-0 | Yes | Yes | 66 |
| NSC 607097 | HIF-1 transcriptional activity and protein accumulation | 497181-30-3 | Yes | No | 70 |
![]()
Inhibitors of HIFα mRNA expression
Although the function of HIFs is controlled primarily at protein levels, HIFα mRNA is a limiting factor affecting the rate of protein synthesis.56 We have previously reported that antisense HIF-1α downregulated the expression of HIF-1α, and enhanced tumor immunotherapy.57 EZN-2968 is a synthetic antisense oligodeoxynucleotide comprised of 16 nucleotide residues which are 100% complementary to residues in the mRNA sequence of human HIF-1α.31 Administration of EZN-2968 downregulated the expression of HIF-1α protein in a dose-dependent manner. It has a three base pair mismatch with the HIF-2α sequence and, as a consequence, showed a weak inhibitory effect on HIF-2α expression. ENZ-2968 was evaluated in phase I clinical trials (NCT01120288, NCT00466583), and the results from one trial showed that administration of ENZ-2968 reduced HIF-1α expression in paired tumor biopsies from 4 out of 6 patients, and the expression of target genes in biopsies from 2 patients. Although the trial provided preliminary proof, it was terminated prematurely due to suspended development of the compound.30 A third phase I trial is underway for hepatocellular carcinoma patients (NCT02564614).
Topoisomerase 1 inhibitors, including irinotecan, topotecan and camptothecin, have been approved for the treatment of various types of cancer. They were also shown to inhibit the expression of HIF-1α.2658 As a result, topotecan is under evaluation in a pilot clinical trial for treating refractory advanced solid neoplasms expressing HIF-1α (NCT00117013). PX-478, a compound derived from melphalan, reduced the expression of HIF-1α mRNA and protein in human tumor xenografts.34 A phase I trial was done with PX-748 for advanced solid tumors and lymphomas, but no results have been reported (NCT00522652).
Inhibitors of HIFα protein synthesis
Although no agents have been developed to selectively inhibit HIFα protein synthesis, several natural agents have been shown to suppress the synthesis of HIFα proteins via unknown or indirect mechanisms. Chrysin, a natural flavonoid, inhibited HIF-1α protein synthesis through AKT signaling,59 while glyceollins, a group of phytoalexins isolated from soybean, potently reduced HIF-1α synthesis by blocking the PI3K/AKT/mTOR pathway.38 2-methoxyestradiol (2ME2) is a product of estrogen metabolite and naturally produced by catechol-O-methyltransferase-mediated O-methylation of 2-hydroxyestradiol. 2ME2 was initially considered to disrupt microtubules, and later found to inhibit synthesis of HIF-1α and HIF-2α proteins, and suppress their nuclear translocation and transcriptional activity.3536 In ongoing phase II trials, 2ME2 is administered alone or in combination with other agents to treat various advanced or metastatic solid tumors (NCT00592579, NCT00481455, and NCT0044431). In addition, KC7F2, a lead compound with a central structure of cystamine, markedly inhibited HIF-1α protein synthesis.37
Inhibitors of protein stabilization and accumulation
Hsp90 is cellular chaperone, which is essential for the folding and localization of target proteins.21 Several Hsp90 inhibitors have been shown to promote HIFα degradation in a VHL-independent manner, such as geldanamycin60 and its analogs, 17-AAG (tanespimycin) and 17-DMAG (alvespimycin). The latter two are under evaluation in clinical trials (NCT00088374 and NCT00089271). Bisphenol A, an endocrine-disrupting chemical, promoted the degradation of HIF-1α protein by dissociating Hsp90 from HIF-1α.42 Recently, EC154, a small molecule Hsp90 inhibitor, was shown to effectively promote degradation of HIF-1α and HIF-2α.61
Histone deacetylase (HDAC) can stabilize HIFα proteins by removing the acetyl groups. Several clinically used HDAC inhibitors, such as vorinostat, have been demonstrated to promote the degradation of HIFα proteins, even though the exact mechanism is not clearly understood.2662
A novel HIF-1α inhibitor (CAS No. 934593-90-5) was shown to reduce the accumulation and transcriptional activity of HIF-1α protein.47 A compound named PX-12 (1-methylpropyl 2-imidazolyl disulfide) inhibited HIF-1α protein accumulation by targeting thioredoxin-1.4849 LW6 is a small molecule that was shown to inhibit the accumulation of HIF-1α via VHL-mediated proteasomal degradation.46 A novel lead structure, BAY 87-2243, identified through high-throughput screenings, was found to suppress the accumulation of HIF-1α and HIF-2α proteins in non-small cell lung cancer cells by inhibiting mitochondrial complex I activity.43 However, a phase I clinical trial for BAY 87-2243 (NCT01297530) was terminated due to safety reasons.
Inhibitors of dimerization
A cyclic peptide inhibitor (cyclo-CLLFVY) was reported to disrupt HIF-1 dimerization and inhibit its transcription activity by binding to the PAS-B domain of HIF-1α. This inhibitor reduced HIF-1-mediated hypoxia response signaling in a variety of cancer cell lines, without affecting the function of HIF-2 isoform.50 On the other hand, two compounds, TC-S7009 and PT2385, were identified to selectively antagonize HIF-2 heterodimerization and DNA-binding, but had no effect on HIF-1 function.5152 PT2385 is under clinical investigation for advanced clear cell renal cell carcinoma in a phase I trial (NCT02293980). Acriflavin, an antiseptic dye and formerly used as a topical antiseptic, was found to inhibit the dimerization of both HIF-1 and HIF-2.53
Inhibitors of DNA binding
Doxorubicin and daunorubicin, two anthracyclines used as conventional chemotherapeutic agents, exert their cytotoxic activity through DNA intercalation. These drugs also effectively inhibited the binding of HIF-1 to the HRE sequences of the target genes.63 Echinomycin (quinomycin A), a small molecule known to bind DNA in a sequence-specific fashion, was shown to inhibit the binding of HIF-1 to the core HRE sequences (5'-CGTG-3') contained in the VEGF promoter.5455
Inhibitors of transcriptional activity
Chetomin, a dithiodiketopiperazine metabolite from the fungus Chaetomium, was found to inhibit the interaction of HIF-1α with p300 by disrupting the tertiary structure of the CH1 domain of p300 protein. Administration of chetomin suppressed the growth of xenografts in mice.64 Indenopyrazole 21, a compound containing the indenopyrazole framework, was found to strongly inhibit HIF-1α transcriptional activity without affecting HIF-1α protein accumulation and HIF-1 heterodimerization.65 YC-1, an antiplatelet aggregation agent, was shown to dissociate the binding of HIFα T-CAD to p300 co-activator, leading to the repression of HIF transcriptional activity; YC-1 could also impair HIFα protein accumulation.2666 FM19G11, a novel chemical entity, significantly repressed p300, a histone acetyltransferase required as a co-factor for HIF-transcription activation, by inhibiting histone acetylation.5567
Go to :
CONCLUSION AND PERSPECTIVES
Hypoxic microenvironments are frequently observed in solid tumors and contribute to the resistance to cancer therapies, including conventional chemotherapy and radiotherapy, and newly developed molecular targeting therapy and immunotherapy.28 The activated HIF pathway, regulated mainly by HIF-1 and HIF-2, is responsible for more malignant phenotypes, aggressive cancer behaviors and poor prognosis. In the past two decades, efforts have been devoted to seek or develop HIF inhibitors. A few inhibitors interfere directly with mRNA or protein of HIF-1 and/or HIF-2, the dimerization of α and β subunits, or the interaction of HIF with its co-activators; but most of them are indirect inhibitors or have multiple activities. Thus, developing more specific HIF inhibitors is still a major challenge. Recent advances in computational molecular modeling technology and high-throughput screening may promote the discovery of new molecular structures and optimization of existing compounds. Some HIF inhibitors have been clinically trialledfor treating solid tumors. Unfortunately, up to now, no drugs directly inhibiting HIFs have been approved for treating cancer patients due to safety or limited therapeutic efficacy.
HIF-1 and HIF-2 share a high degree of sequence identity, and both contribute to the regulation of cellular adaption to hypoxia. However, little is known how they cooperate or compete with each other. HIF-1α plays the dominant role in the response to acute hypoxia, whereas HIF-2α drives the response to chronic hypoxia, although both are involved in cancer progression.18 Importantly, depletion of one HIF α-subunit provokes a compensatory increase of the other one, such that knockdown of HIF-1α increases the expression of HIF-2α via the reciprocal regulatory mechanism, and this switch from HIF-1α- to HIF-2α-dependent ways provides a mechanism for more aggressive growth of tumors and drug resistance.176869 Interestingly, these studies have provided a clue that it may be essential to target HIF-1 and HIF-2 together, rather than one factor, so as to suppress hypoxia-inducible pathways. The majority of current HIF inhibitors, however, are targeting HIF-1. Therefore, developing HIF-2 inhibitors and agents that are able to simultaneously block HIF-1 and HIF-2 may provide more effective approaches for cancer treatments.
Little is known about HIF-3, the newest member of HIF family, compared with the other two homologues. However, a few recent studies indicate that HIF-3α have multiple variants due to different promoters, transcription initiation sites and alternative splicing. Some HIF-3α variants are dominant-negative regulators of HIF-1/2α, while others inhibit HIF-1/2α actions by competing for the common β-subunit.13 Therefore, further studies on the HIF-3α variants may provide new insights into the HIF biology.
Because of the rapid degradable feature of α subunits of HIF under normoxia, the HIF pathway is activated in hypoxic cancer cells or tissues. Contradictorily, hypoxic areas in solid tumors are caused mainly by deteriorating diffusion geometry, which could also prevent the delivery of HIF inhibitors to reach hypoxic cancer cells. Therefore, enhancing the penetrating capacity of HIF inhibitors from capillaries to cells may strengthen their anti-cancer effects.
The lack of patient selection also contributes to the failure of clinical trials. In most of the clinical trials for evaluating the efficacy of HIF inhibitors, patients with advanced or refractory solid tumors are recruited, regardless of the expression levels of HIF in tumors. In this case, HIF inhibitors may not display therapeutic effects for those that have low levels of HIF expression. Therefore, the concept of personalized medicine should be applied in designing clinical trials for HIF inhibitors, so that they may be more effective for a specific subpopulation of patients whose tumors have elevated HIF levels.
Go to :
 XML Download
XML Download