

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cardiovascular diseases, such as coronary artery disease, atherosclerosis, congestive heart failure, hypertension, stroke, and myocardial infarction, are a major cause of death in the world.123 Studies have demonstrated that abnormal proliferation of vascular smooth muscle cells (VSMCs) play a critical role in the pathogenesis of cardiovascular diseases.456 However, to our best knowledge, the underlying molecular mechanisms thereof remain unclear.
Platelet-derived growth factor-bb (PDGF-bb) is released primarily by vascular endothelial cells and platelets at the sites of vascular injury, and it has been shown to potently stimulate VSMCs proliferation and migration by modulating several key molecular signaling pathways.78 Several studies have demonstrated that increased expression of signaling proteins in the PDGF-bb pathway is associated with cardiovascular diseases, including restenosis and atherosclerosis,9 and inhibition of PDGF-bb-related pathways has been shown to exert beneficial effects on these cardiovascular disorders.10 However, the underlying mechanisms by which PDGF-bb modulates VSMC proliferation and migration are not fully understood. Therefore, it is of great scientific interest to explore the molecular mechanisms involved in the modulation of PDGF-bb-dependent VSMC proliferation and migration.
MicroRNAs (miRNAs) are a class of small non-coding RNAs, by targeting the 3' untranslated region (3'UTR) of mRNA, miRNAs can induce mRNA degradation, which in turn results in the translation repression of target genes.11 Various studies have showed that miRNAs are important in cell development, growth, differentiation, proliferation, and apoptosis.121314 Increasing evidence has also demonstrated that aberrant expression of miRNAs is involved in the development and progression of many types of cancers, including colon cancer, liver cancer, gastric cancer, lung cancer, breast cancer, and so on.151617 Meanwhile, recent studies have found that miRNAs also regulate the functions of VSMCs: for example, Xu, et al.18, demonstrated that miR-135b and miR-499a promote cell proliferation and migration in the atherosclerosis by directly targeting myocyte enhance factor 2C; Liu, et al.19, reported that miR-1 regulates the proliferation of VSMCs by targeting insulin-like growth factor-1 (IGF-1); and Xie, et al.20, showed that miR-599 inhibits VSMCs proliferation and migration by targeting transforming growth factor beta 2.20 However, the role of miRNAs in VSMC functions has not been fully explored.
Xu, et al.18, reported that miR-379 was down-regulated in circulating blood from patients with atherosclerotic coronary artery disease. miR-379 has also been found to inhibit cell growth in certain types of cancers including breast cancer, malignant mesothelioma, and liver cancer.212223 In this study, we investigated the functional role of miR-379 in the growth of VSMCs. Herein, PDGF-bb treatment promoted proliferation of VSMCs and down-regulated the expression of miR-379. Functional assays demonstrated that miR-379 inhibits cell proliferation, cell invasion, and migration. Flow cytometry results showed that miR-379 induces apoptosis in VSMCs. Using a bioinformatic analytic tool (Targetscan; http://www.targetscan.org), the 3'UTR of IGF-1 gene was found to be a target of miR-379. Luciferase report assay further confirmed that IGF-1 3'UTR is a direct target of miR-379, and the expression of miR-379 and IGF-1 at both mRNA and protein levels were inversely correlated. Finally, rescue experiments showed that enforced expression of IGF-1 sufficiently overcomes the inhibitory effect of miR-379 on cell proliferation in VSMCs.
MATERIALS AND METHODS
Cell culture
VSMC lines (Cell bank of Chinese Academy of Science, Shanghai, China) were cultured in Dulbecco's modified Eagle's medium (DMEM) (Hyclone, GE Health Care, Chicago, IL, USA) supplemented with 10% fetal bovine serum (FBS). VSMCs were stored in an incubator with 5% CO2 at 37℃.
miRNAs and plasmids
miR-379 mimics, miR-379 inhibitors, and scrambled miRNA were purchased from GenePharma (Shanghai, China). The empty vectors pcDNA3.1 and pcDNA3.1-IGF-1 were synthesized by Ribobio (Guangzhou, China).
Transfection and PDGF-bb treatment
Lipofectamine 2000 (Invitrogen, Waltham, MA, USA) was used to perform miRNA transfection according to the manufacturer's instructions. Co-transfection of miR-379 mimics and pcDNA-3.1-IGF-1 were performed according to the similar procedures above. PDGF-bb (Sigma, St. Louis, MO, USA) treatments were carried at a concentration of 10–30 ng/mL, and the CCK-8 assay were performed at 24, 48, and 72 h.
Preparation of RNA and quantitative real-time PCR
Total RNAs were extracted using Trizol reagent (Invitrogen). The TaqMan MicroRNA Reverse Transcription Kit (Takara, Dalian, China) and the TaqMan Reverse Transcription Kit (Takara) were used to reverse transcribe RNA for miRNA detection and to obtain cDNA for mRNA detection, respectively. For miR-379 and IGF-1 mRNA, quantitative real-time PCR (qRT-PCR) was performed using miRscript SYBR Green PCR Kit and SYBR Green PCR Kit (Takara), respectively. The internal controls for miR-379 and IGF-1 mRNA were U6 and GAPDH, respectively. The primers for miR-379 were forward,5'-GCTACATGATAC AGTGCAAA-3' and reverse, 5'-AGTTTGCTTGATCCCTCTTC AG-3'; the primers for IGF-1 mRNA were forward, 5'-CTTCA GTTC GTGTGT GGA GAC AG-3' and reverse, 5'-CGCC CTCC GACTGCT-3'. Data are expressed as fold changes normalized to U6 or GAPDH based on the following formula: RQ=2-ΔΔCt.
Western blot
Proteins extraction was performed using RIPA lysis buffer (Bio-Rad, Milan, Italy). The extracted protein lysates were separated by sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and then transferred to a polyvinylidene difluoride membrane (PVDF, Millipore, Billerica, MA, USA). Membranes were blocked with 5% bovine serum albumin, followed by overnight incubation with primary antibody at 4℃: rabbit anit-IGF-1 (1:2500; Abcam, Cambrigde, MA, USA) and mouse anti-β-actin (1: 7000; Abcam, Cambridge, MA, USA). Membranes were then incubated with horseradish peroxidase-conjugated secondary antibodies (1:4000; Abcam). The membranes were exposed using a ChemoDoc XRS detection system (Bio-Rad, Milan, Italy).
Cell proliferation assay
For the CCK-8 assay, cells were seeded in 96-well plates (5×103 cells/well), and incubated for 24 h. Cells were then treated with PDGF-bb; transfected with miR-379 mimics, miR-379 inhibitor, or their respective controls; or co-transfected with miR-379 and pcDNA3.1-IGF-1. At 24, 48, and 72 h after PDGF-bb treatment or at 48 h after transfection, cell proliferation indices were measured using a CCK-8 kit (Beyotime, Beijing, China).
Cell invasion assay
For invasion assay, cells were seeded on 24-well transwell plates (5×104 cells/well), cultured for 24 h, and then transfected with miR-379 mimics, miR-379 inhibitor, or their respective controls. Cells were grown in the upper chambers of the plates in serum-free DMEM separated by a Matrigel-coated membrane (BD Bioscience, San Jose, CA, USA), and 500 µL of DMEM supplemented with 10% FBS was added to the bottom of the chambers. Cells were then incubated for 24 h at 37℃, and were stained with 0.1% crystal violet.
Wound healing assay
For the wound healing assay, cells were seeded in six-well plates (5×105 cells/well). Twenty-four hours after transfection, a scratched wound was created using a pipette tip. Cells migrated into the wound surface, and the average distance of the migrating cells was determined under an inverted microscopy at 0 h and 24 h.
Luciferase reporter assay
The constructed wild type or mutant miR-379 binding site of IGF-1 3'UTR pmirGLO vectors were purchased from Ribobio (Guangzhou, China). VSMCs were cultured on 24-well plates 24 h before co-transfected with 50 ng of wild type or mutant luciferase vector and 20 µM miR-379 mimics, miR-379 inhibitor, or their respective controls. After 48 h, luciferase activity was measured by the Dual-luciferase Reporter Assay System (Promega, Madison, WI, USA).
Cell apoptosis assay
For cell apoptosis analysis, cells were seeded on a six-well plate, and 24 h post-transfection, cells were harvested and washed with phosphate buffered saline (PBS). Cell apoptosis was then detected using an Annexin V-FITC/PI apoptosis detection kit according to the manufacturer's instructions.
Statistical analysis
GraphPad Prism version 6 (GraphPad Prism version 6.0, Inc., La Jolla, CA, USA) was used to perform all statistical analyses. Significant differences between groups were determined by t-test or one-way ANOVA followed by Bonferroni's multiple comparison tests. All data are expressed as the mean values of three independent replicates±SEM; differences were considered significant when p<0.05.
RESULTS
PDGF-bb promotes cell proliferation and inhibits miR-379 expression in VSMCs
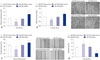
The effect of PDGF-bb on cell proliferation of VSMCs was assessed by CCK-8 assay. As shown in Fig. 1A and B, PDGF-bb promoted proliferation of VSMCs in a concentration-dependent and time-dependent manner. The expression of miR-379 in VSMCs was determined by qRT-PCR, and PDGF-bb treatment significantly suppressed the expression level of miR-379 in VSMCs (Fig. 1C and D).
miR-379 inhibits cell proliferation, invasion and migration of VSMCs
To validate the inhibitory role of miR-379 on proliferation of VSMCs, cell proliferation assay was performed by using CCK-8 kit. Compared to their respective control groups, the proliferative ability of miR-379 mimics-transfected VSMCs were significantly inhibited, while the proliferative ability of miR-379 inhibitor-transfected VSMCs were significantly enhanced (Fig. 2A). Cell invasion assay was performed to further examine if the differential expression of miR-379 was correlated with cell invasion. As shown in Fig. 2B, miR-379 mimics transfection in VSMCs caused lower invasion ability when compared to that transfected with control; meanwhile, miR-379 inhibitor transfection enhanced the invasive ability of VSMCs (Fig. 2B). Wound healing assay was conducted to determine the effect of miR-379 on the motility of VSMCs. Transfection of miR-379 mimics in VSMCs caused a decrease in wound healing cell migration, compared to its control group; transfection of miR-379 inhibitor in VSMCs increased wound healing cell migration, compared to its control group (Fig. 2C).
miR-379 induced cell apoptosis in VSMCs
To examine if miR-379 induces apoptosis in VSMCs, we further performed flow cytometry experiment. As shown in Fig. 2D, transfection with miR-379 mimics increased the cell apoptotic rate in VSMCs, compared to its respective control, while miR-379 inhibitor treatment caused a decrease in the cell apoptotic rate of VSMCs, compared to its respective control (Fig. 2D).
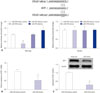
IGF-1 is a direct target of miR-379
The downstream mediator of miR-379 was predicted by TargetScan (Supplementary Table 1, only online), and there were complementarities between miR-379 and IGF-1 3'UTR (Fig. 3A). Co-transfection with miR-379 mimics and the wild type reporter plasmid significantly suppressed the relative luciferase activity of the reporter containing wild type IGF-1 3'UTR. Co-transfection of miR-379 inhibitor and the report plasmid significantly increased the luciferase activity of the reporter containing wild type IGF-1 3'UTR, while the luciferase activity of the reporter containing mutant IGF-1 3'UTR was unaffected (Fig. 3A). Further, qRT-PCR and western blot results showed that the expressions of miR-379 and IGF-1 at both the mRNA and protein level were inversely correlated (Fig. 3B and C). To further investigate the role of IGF-1 in cell proliferation, invasion, and migration of VSMCs, pcDNA3.1-IGF-1 and miR-379 mimics were co-transfected into VSMCs, and CCK-8 assay, invasion assay, and wound healing assay were performed, respectively. As shown in Fig. 4, enforced expression of IGF-1 was able to sufficiently overcome the inhibitory effect of miR-379 on the proliferation, invasion, and migration of VSMCs (Fig. 4).
DISCUSSION
In the present study, we observed that PDGF-bb promotes VSMCs proliferation, and the expression of miR-379 was suppressed in VSMCs by PDGF-bb treatment. Functional assays confirmed that miR-379 inhibited cell proliferation, cell invasion, and migration. Flow cytometry results demonstrated that miR-379 induces apoptosis in VSMCs. Reporter gene assay, qRT-PCR, and western blot showed that translation suppressed by miR-379 was mediated by IGF-1 mRNA 3'UTR dependent on the presence of a single miR-379 cognate binding site, and enforced expression of IGF-1 overcame the inhibitory effect of miR-379 on the proliferation, invasion, and migration of VSMCs. The above results suggest that miR-379 plays a crucial role in controlling VSMC proliferation.
miRNAs have been implicated in various cell functions, including differentiation, proliferation, death, metabolism, and carcinogenesis.121314 Deregulation of miRNAs has been found in many types of human cancers.151617 Recent studies have found that miRNAs also regulated the functions of VSMSCs.181920 As PDGF-bb treatment promotes VSMC proliferation and migration, changes in miRNA expression profiles by PDGF-bb may be of great interest to identify novel miRNAs, which may regulate proliferation and migration. In our preliminary studies, we treated VSMCs with PDGF-bb and examined the expression levels of several miRNAs potentially involved in the regulation of VSMCs (Supplementary Table 2, only online). The preliminary results showed that miR-379 is down-regulated by PDGF-bb stimulation (Supplementary Table 2, only online); therefore, miR-379 was chosen for further functional study. To date, the role of miR-379 in VSMC function has not been investigated. One study found that miR-379 levels are lower in circulating blood from patients with atherosclerotic coronary artery disease.18 In cancer studies, miR-379 was found to inhibit breast cancer cell proliferation by regulating cyclin B1 expression;21 Yamamoto reported that miR-379 inhibited cell growth of malignant pleural mesothelioma by targeting interleukin 18.23 In addition, miR-379 also exhibited a tumor-suppressive function in liver cancer.22 Consistently, our results showed that miR-379 inhibited cell proliferation, cell invasion, and migration of VSMCs.
IGF-1 is a growth factor and has been found to regulate cell proliferation and migration of VSMCs.24 Studies have demonstrated that over-expression of IGF-1 promotes neointimal formation after carotid artery injury, which results from an increase in the proliferation and migration of VSMCs.25 IGF-1 exerts its functions by acting on IGF-1 receptor, which is expressed in VSMCs.26 In the present study, the 3'UTR of IGF-1 gene was a target of miR-379, as predicted by TargetScan, and this predicted target was further confirmed by luciferase report assay. Furthermore, miR-379 and IGF-1 expressions at both the mRNA and protein level were inversely correlated. In addition, rescue experiment showed that enforced expression of IGF-1 overcame the inhibitory effect of miR-379 on cell proliferation of VSMCs. Collectively, these results may suggest that IGF-1 potentiates the proliferation of VSMCs, which can be suppressed by overexpression of miR-379. It has been demonstrated that miRNA may target more than one gene; it is likely that miR-379 targets both IGF-1 and other genes to regulate cell proliferation, invasion, and migration of VSMCs. However, in the present study, we did not show whether miR-379 targets other genes (Supplementary Table 1, only online) that modulate cell proliferation, migration, and invasion, and thus, further study may be required to confirm the role of other genes targeted by miR-379 in VSMCs.
In conclusion, the present study indicates that miR-379 plays a key role in regulating VSMCs proliferation, invasion, and migration via targeting IGF-1.


 XML Download
XML Download