

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Intracranial aneurysm (IA) accounts for approximately 85% of all subarachnoid hemorrhages (SAHs), which have a high mortality rate of up to 50%.1 Extracellular matrix (ECM) provides structural integrity to the arterial wall, and disruption of the ECM may be related to IA formation.2 Lysyl oxidase (LOX) is a copper-containing amine oxidase that regulates cross-linking and maturation of collagen and elastin.34 Accordingly, the LOX gene has been investigated as a candidate for IA development.4 Mäki, et al.5 reported that inactivation of the LOX gene led to aortic aneurysm caused by structural alteration in the arterial wall in mice. Onda, et al.6 found three linkage regions of IA, chromosome 5q22-31, 7q11 and 14q22, in Japanese sib pairs.6 However, LOX mapped to 5q31 did not show allelic and haplotype-based association with IA in the Japanese population. In addition, no significant associations of LOX variants with IA were observed in a South Indian population.7 Nevertheless, LOX has been reported to be a risk factor for coronary artery disease (CAD) and cerebral stroke. Ma, et al.8 found that G473A polymorphism of LOX was associated with CAD. Zhang, et al.9 reported that frequencies of LOX 473AA genotype and an A allele were significantly higher in patients with ischemic stroke, compared to controls, in a Chinese population. A missense mutation of LOX (c.893T>G encoding p.Met298Arg) was also associated with thoracic aortic aneurysm in humans.10 Such data suggest that LOX variants could be related to IA formation by degenerative endothelial remodeling due to insufficient cross-linking of elastin and collagen fibers in the region of high wall share stress or high spatial pressure gradients.1112
To the best of our knowledge, up to now, a genetic association study of LOX gene polymorphisms has not been conducted in Koreans. Therefore, in an attempt to assess genetic risks in affected IA patients, we aimed to identify LOX gene polymorphisms giving rise to susceptibility to IA in a homogeneous Korean population.
MATERIALS AND METHODS
Study population
This prospective study included 80 radiologically confirmed IA patients with saccular shape, as well as 80 age-and sex-matched controls, from March to December 2016 at a single institution. Non-saccular aneurysms featuring fusiform or dissection and traumatic or infectious aneurysms were excluded. The control group consisted of matched patients who underwent computed tomography or magnetic resonance angiography for headache evaluation or a medical check-up, excluding other neurological diseases, such as arteriovenous malformation, intracranial hemorrhage or infarct, etc. Medical records were reviewed concerning multiple variables, such as sex, age, clinical presentation such as unruptured IA or SAH, hypertension (HTN), diabetes mellitus (DM), hyperlipidemia, smoking,13 and familial history of aneurysm (vs. sporadic). Angiographic variables were reviewed regarding the size of the aneurysm, location (anterior vs. posterior circulation),14 and number (single vs. multiple). This study was approved by the Institutional Review Boards (No. 2016-31).
SNP selection and genotyping
We selected 10 tagging single nucleotide polymorphisms (SNPs) located between 20 kb 5′-upstream and 3′-downstream of the LOX gene after applying a linkage disequilibrium (LD; r2<0.8) in Japanese and Chinese (JPT+CHB) HapMap database (Phase II) from the LD TAG SNP Selection (TagSNP) of SNPinfo (https://snpinfo.niehs.nih.gov/). For genotyping of 10-tagged SNPs, genomic DNA from the peripheral blood of all 160 subjects was extracted using HiGene™ Genomic DNA Prep Kit (BIOFACT, Daejeon, Korea). Primers of ten SNPs were designed using the Primer-3 v.0.4.0 program (http://bioinfo.ut.ee/primer3-0.4.0/) (Table 1). Polymerase chain reaction (PCR) was performed at 25-uL volume containing 100 ng genomic DNA with 1.5 uL per primer (10 pmole/uL) by Solg™ 2X Taq PCR Pre-Mix (Solgent, Daejeon, Korea). Pre-denaturation was done at 95℃ for 5 minutes, 34 cycles of denaturation at 95℃ for 30 seconds, annealing at 63℃ for 30 seconds, extension at 72℃ for 1 minute, and a final extension at 72℃ for 5 minutes. The amplified fragments were confirmed by 1.5% agarose gel electrophoresis, purified with the Solg™ PCR purification kit (SolGent, Daejeon, Korea), and sequenced by the ABI PRISM 3730XL Analyzer (Applied Biosystems, Foster City, CA, USA).
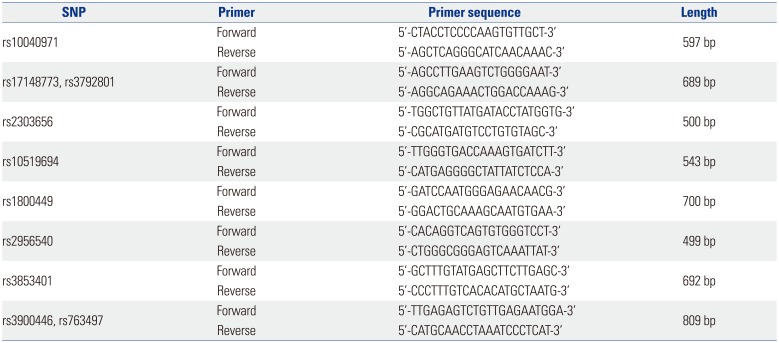
Table 1
Primers Designed for 10 SNPs of the Lysyl Oxidase Gene
![]()
Statistical analyses
Continuous data are expressed as means±standard deviation. Kruskal-Wallis test with descriptive analysis was conducted to evaluate the difference between 80 IA patients and 80 controls for non-genetic factors. Fisher's exact test was performed to assess allelic associations between IA and ten SNPs located near or on the LOX gene to estimate odds ratios (ORs). In subsequent analysis, we analyzed the genetic effects of the LOX gene on IA formation according to aneurysm rupture. Furthermore, an omnibus test with haplotype-specific association analysis, which contains h-1 degrees of freedom (h, the number of all possible haplotypes in a sliding window depending on the number of SNPs), was conducted using asymptotic chi-square statistics to identify significant haplotype associations with IA. This test excluded haplotype structures with a minor haplotype frequency (MHF) less than 0.01 of possible combinations using ten SNPs. We applied Bonferroni-adjusted significant p value less than 0.005 and 2.0×10−4 after testing multiple comparison corrections and suggestive significant p values of 0.01 and 0.001 in single SNPs and haplotype associations, respectively. The descriptive and univariate analyses were performed by STATA software v.11.2 (Stata Corp., College Station, TX, USA). Quality control test for 10 SNPs was conducted to evaluate the genotyping call rate (GCR), minor allele frequency (MAF), Hardy Weinberg equilibrium (HWE) p value, and pairwise LD using Haploview v.4.2 (https://www.broadinstitute.org/haploview/haploview).15 Genetic and haplotype associations were analyzed using PLINK program v.1.07 (http://zzz.bwh.harvard.edu/plink/).16
RESULTS
Demographic characteristics of the enrolled patients
The baseline characteristics of the two groups (IA and controls) are described in Table 2 and Supplementary Table 1 (only online). In the IA group, the number of females was 43 (53.8%), and their mean age was 57.1±12.9 years. SAH presentation was noted for 41 (51.3%) patients. Regarding aneurysm location, anterior circulation aneurysms (n=73, 91.3%) were noted as follows: internal carotid artery, n=15; anterior communicating artery or anterior cerebral artery, n=21; middle cerebral artery, n=25; and posterior communicating artery, n=12. Seventy-five (93.8%) patients had a single aneurysm. No patient who had a familial history of SAH was observed in our study. Between the two groups, the incidences of HTN, DM, hyperlipidemia, and smoking did not differ significantly.
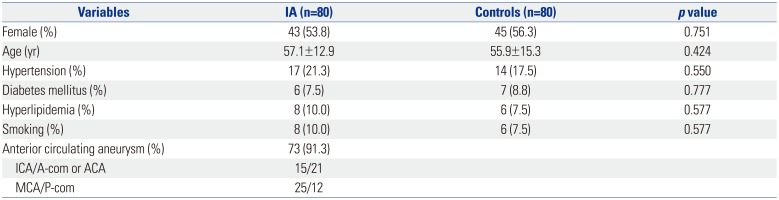
Table 2
Baseline Characteristics of Patients with IA and Controls
![]()
Genetic associations of 10 LOX gene polymorphisms with IA
All ten SNPs located near or on the LOX gene showed completed GCR, MAF greater than 0.01, HWE p-value greater than 0.05, and LD greater than 0.8 after quality control tests. The LD structures of ten SNPs in patients with IA, the control group, and all subjects are shown in Fig. 1. There remained LD in rs10040971 with rs3792801 (r2=1.00) and rs10519694 with rs3853401 (r2=0.89) in patients with IA and rs2956540 with rs1800449 in controls. Among ten SNPs, three SNPs, rs2303656, rs3900446, and rs763497, showed statistically significant associations with IA (p<0.05) (Table 3) (Supplementary Table 2, only online). Two SNPs, rs2303656 and rs3900446, reached the Bonferroni-adjusted significance threshold (p<0.005). The C allele of rs3900446 showed the most significant and strongest associations with an increased risk of IA (OR=20.15, p=4.8×10−5). On the contrary, the A allele of rs2303656 showed a protective effect on IA and was not frequently observed in the patient group (p=8.2×10−4). The G allele of rs763497 showed a suggestive association with an increased IA risk (OR=2.26, p=0.009). However, none of the SNPs was associated with IA rupture among 80 patients with IA in subsequent analysis (p>0.05).
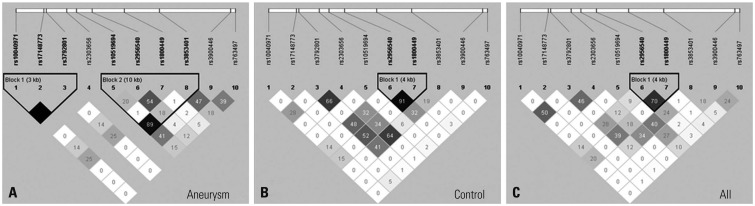
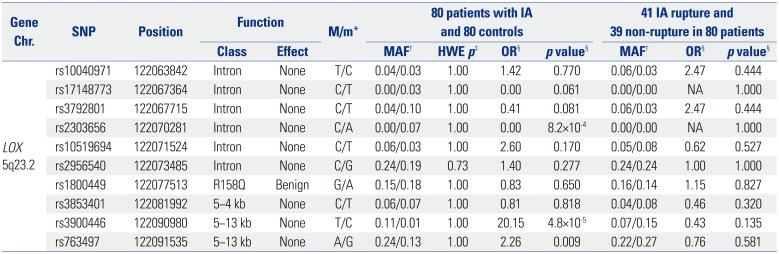
Fig. 1
Linkage disequilibrium patterns of 10 Lysyl oxidase SNPs in patients with intracranial aneurysm (A), controls (B), and total groups (C). SNPs, single nucleotide polymorphisms.
![]()
Table 3
Results of 10 SNPs Associations with IA
Chr., chromosome; IA, intracranial aneurysm; NA, not available; OR, odds ratio; MAF, minor allele frequency; HWE, Hardy-Weinberg equilibrium; SNPs, single nucleotide polymorphisms.
*Major/minor allele type, †MAF in case (left) and control (right) groups, ‡HWE p-value for control group, §OR and p value were estimated from allelic association analysis using Fisher's exact test.
![]()
In an omnibus test of haplotype association, 136 of total 247 haplotype structures in 45 sliding windows (SNP set) showed an asymptotic p-value less than 0.05 (data not shown), and 17 haplotypes showed a suggestive association with IA (asymptotic p<0.001) (Table 4). Of ten SNP haplotype combinations, the CG combination of rs3900446 and rs763497 showed significant associations in a single SNP analysis (MHF=0.113, asymptotic p=1.3×10−5) (Table 3). Although six SNPs (rs10040971, rs17148773, rs3792801, rs10519694, rs2956540, and rs1800449) were not independently associated in single SNP analyses, the haplotype structures of combining these SNPs with either rs2303656, rs3900446, or rs763497 were significant in haplotype analyses (6.5×10−4<p<7.5×10−4) (Table 4).
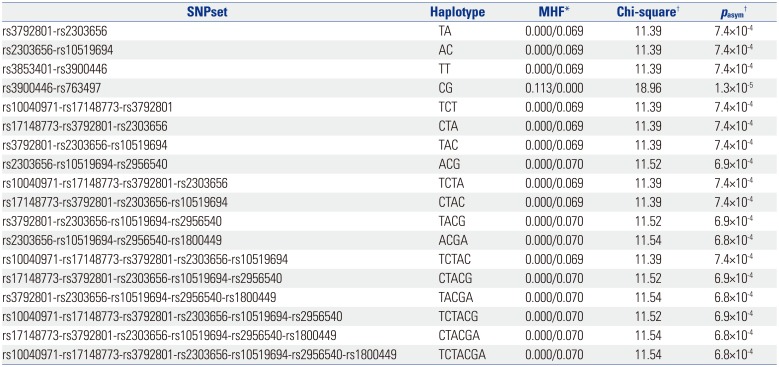
Table 4
Results of Haplotype Associations with Intracranial Aneurysm
![]()
DISCUSSION
Our results showed that three SNPs (rs2303656, rs3900446, and rs763497) were significantly associated with IA in a Korean population. The T allele of rs3900446 was significantly related to an increased IA risk with a significant threshold (OR=20.15, p=4.8×10−5). On the contrary, the A allele of rs2303656 revealed the protective effect on IA formation. Haplotype analysis showed that the CG combination of rs3900446 and rs763497 reached Bonferroni-adjusted significant threshold in IA patients (MHF=0.113, asymptotic p=1.3×10−5).
Aneurysms featured multifactorial disorder affected by environmental and genomic factors. Variables, such as HTN, smoking and larger size at diagnosis, have been reported to be associated with IA formation and growth.17 Subjects with first-degree IA relatives showed four times higher incidence of aneurysm, compared to those without IA relatives.18 Compared to sporadic aneurysms, familial aneurysms showed larger size of aneurysm at the time of rupture and multiplicity.19 After reviewing 10 genome-wide linkage analyses of familial IA, four loci, such as 1p34.3-p36.13, 7q11, 19q13.3, and Xp22, were demonstrated in other cohorts.20 Positional and functional candidate genes of elastin and collagen type 1 A2 were associated with 7q11 and perlecan with 1p34.3-p36.13. Regarding sporadic IA, SNPs on chromosome 4 near the endothelin receptor A gene (rs6841581), chromosome 9 within the cyclin-dependent kinase inhibitor 2B antisense inhibitor gene (rs10757278 and rs1333040), and chromosome 8 near the SOX17 transcription regulator gene (rs9298506 and rs10958409) were significantly associated with aneurysm.21 Beyond the suggestive loci for IA as mentioned above, versican (VCAN) gene located at the locus of 5q22-31 has been reported to be related to IA. Two SNPs rs251124 and rs173686 in strong LD and haplotypes were associated with IA in a Dutch population.22 Sathyan, et al.23 reported that rs251124 was the strongest marker of IA for global ethnicities. In addition, a novel association of IA with rs2287926 (G428D) in exon 7 coding has been reported. Other potential candidate genes, such as LOX, fibroblast growth factor 1 (FGF1) and fibrillin 2 (FBN2), can be included in the linkage region of 5q22-31. Yoneyama, et al.24 reported difference in allelic frequency for SNP at intron 4 of FGF1 (χ2=4.44, df=1, p=0.035); however, no association of SNP in LOX or FBN2 was noted. Hofer, et al.4 also did not find an allelic association or co-segregation in 25 German IA families after analyzing four genetic variants. Ruigrok, et al.25 analyzed the 44 potential candidate genes of ECM integrity in a Dutch population in developing IA. They reported that serpine 1 (SERPINE 1, combined OR 1.27, p=0.004), FBN2 (combined OR 1.37, p=0.01), and alpha 1 type IV collagen (COL4A1, combined OR 1.22, p=0.007) were related to IA; however, no association of SNPs between the LOX gene and IA was demonstrated. Sathyan, et al.7 did not find allelic or genotypic variants of the LOX gene in a South Indian population with IA. Several genome-wide association studies2627 also did not demonstrate the 5q22-31 loci for IA formation. In contrast, three SNPs (rs2303656, rs3900446, and rs763497) were significantly associated with IA in Koreans in this study. Differences in the frequencies of genetic polymorphisms according to ethnics have been reported. Tian, et al.28 reported the genetic diversity of many SNPs in cancer-related genes between Chinese Hui and Han populations. Inoue, et al.29 also showed a difference in the frequency distribution of the genetic polymorphisms in the CYP1A1 and CYP1B1 gene in Japanese and Caucasian populations. Accordingly, ethnic differences should be carefully considered in evaluating risk factors for the disease.30
Hemodynamic factors have been thought to be related to aneurysm formation. The fluid dynamic study revealed that elevated wall share stress by repetitive flow impingement may contribute to aneurysm formation by degenerative endothelial remodeling.1112 In clinical circumstances, most aneurysms were located at the arterial branching and bifurcation sites. Alterations in the aneurysm wall and, in particular, focal degradation of the ECM have been linked to aneurysm formation and growth.31 Bruno, et al.31 reported that aneurysm tissue showed a higher level of membrane-type matrix metalloproteinase (MMP) and MMP-2, compared to controls. LOX controls the cross-linking and maturation of elastin and collagen fibers, which are responsible for mechanical stability of the arterial wall.4 Accordingly, LOX gene variants could lead to ECM instability and aneurysm formation by inducing insufficient stability to mechanical stimuli on the arterial wall.
There are some limitations in this study. First, the sample size was relatively small, although LOX gene polymorphisms were observed to have strong associations with IA and likely to imply a sufficient statistical power due to its large effect size.32 The second GWAS by Yasuno, et al.33 showed additional three new loci near retinoblastoma binding protein 8 (RBBP8) on 18q11.2, StAR-related lipid transfer domain containing 13/Klotho (STARD13/KL) on 13q13.1, and a gene-rich region on 10q24.32, as well as prior associations near SOX17 and CDKN2A/B. However, the LOX gene was not associated with IA in their study. One possible explanation for missed loci is that two SNPs of rs2303656 and rs3900446 are rare variants in Europeans or Japanese than Koreans. However, small sample size could also affect the high risk of C allele of rs3900446 for IA formation. Second, no replication for the associations between LOX gene polymorphisms and IA was performed. Third, supportive functional data was not provided. Accordingly, our novel findings should be replicated in other populations via further studies including in silico functional studies. Nevertheless, this study is the first genetic association study of LOX gene polymorphisms and IA in Koreans.
In conclusion, this preliminary study showed that three SNPs (rs2303656, rs3900446, and rs763497) were significantly associated with aneurysm formation in Koreans. The C allele of rs3900446 was identified to be strongly associated with an increased risk of IA, while the A allele of rs2303656 showed a protective effect on IA formation. According to haplotype analysis, the CG combination of rs3900446 and rs763497 may contribute to an increased risk of IA. Our findings may play crucial roles in understanding the pathogenesis of IA and provide information on improving genetic risk prediction of IA.
 XML Download
XML Download