

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Gastric cancer is the fourth most common cancer and the second leading cause of cancer-related deaths worldwide.1 Extracellular matrix (ECM) proteins play an important role in tumor progression and metastasis.23 Therefore, a better understanding of ECM proteins present in the tumor microenvironment and their influence on cell-ECM interactions could aid the prevention and treatment of gastric cancer progression and metastasis.
The ECM protein periostin has pathophysiological roles in bone formation,4 wound repair,5 vascular diseases,6 and tumor development,7 and has also been implicated in normal physiological
processes, including cardiac development. It is highly expressed in gastric cancer and is indispensable for successful progression and metastasis.89 Clinical studies have showed that high periostin expression correlates with tumor metastasis and poor prognosis.1011 Periostin is of increasing interest in gastric cancer because it is functionally involved in multiple steps of cancer progression and participates in different signaling pathways. These include metastatic niche formation,12 maintenance of stemness,13 EMT,14 angiogenesis, and the survival of tumor cells,15 all of which are indispensable for gastric cancer progression and metastasis. Periostin also participates in and promotes tumor progression through the FAK/Src, Wnt, and PI-3K/AKT signaling pathways.1617 Therefore, periostin represents a promising candidate for the inhibition of gastric cancer progression and metastasis.
In this study, we investigated the expression of periostin in primary gastric tumors and metastases. First, we examined the expression of periostin in the primary tumours and metastases by in situ hybridization. Next, we evaluated the location of periostin and characteristics of its distribution in gastric cancer using immunofluorescence. Then we investigated the effects of isoprenaline on the expression levels of periostin in gastric cancer cells. Our results demonstrated that periostin is overexpressed by pericryptal fibroblasts in gastric cancer tissues, and were positively correlated with the expression of EMT-associated protein, α-smooth muscle actin (SMA). Finally, we found that the distribution patterns of periostin were broader while the clinical staging of tumor progressed, and isoprenaline upregulated expression levels of periostin in gastric cancer cells.
MATERIALS AND METHODS
Materials
Gastric tissues were collected from the General Hospital of PLA who underwent curative surgical resection with informed consent of patients and the institutional approval. Normal, cancer and metastatic gastric tissue from lymph nodes and tissue adjacent to the tumor were collected from patients diagnosed with gastric cancer. Human gastric adenocarcinoma cell lines MKN-45 and BGC-803 were obtained from the Cancer Institute, Chinese Academy of Medical Science. RPMI-1640 medium, 0.25% trypsin, 0.02% EDTA and fetal bovine serum (FBS) were purchased from Gibco (San Diego, CA, USA); QPCR master mix and the M-MLV reverse transcription system were purchased from Promega (Madison, WI, USA); anti-α-SMA mouse monoclonal antibody and anti-periostin rabbit monoclonal antibody, anti-β-actin mouse monoclonal antibody were purchased from Abcam (Boston, MA, USA); peroxidase-conjugated affinipure goat anti-rabbit IgG, peroxidase-conjugated affinipure goat anti-mouse IgG, Alexa Fluor594-conjugated affinipure goat anti-rabbit IgG and Alexa Fluor488-conjugated affinipure goat anti-mouse IgG were purchased from Cell Signaling Technology (Boston, MA, USA).
In situ hybridization
Antisense and sense cRNA probes were prepared by in vitro transcription. An EcoRI-XbaI fragment of human and mouse periostin cDNA fragment were labeled by digoxygenin using a DIG RNA Labeling Kit (Roche Applied Science, Indianapolis, IN, USA).18 In situ hybridization was performed manually on paraffin-embedded sections (5 mm thick) as described previously.19 The signals were developed using nitroblue tetrazolium salt and 5-bromo-4-chloro-3-indolylphosphate.
Immunofluorescent staining and laser scanning confocal microscopy
Immunohistochemistry was performed as previously described.20 Paraffin-embedded sections of gastric tissues were fixed with cold methanol at 4℃ for 30 minutes, and then permeabilized with 0.2% Triton X-100 in phosphate-buffered saline (PBS) at room temperature for 10 minutes. The sections were stained with anti-periostin antibody (1:100), anti-α-SMA antibody (1:400) and the appropriate Alexa-Fluor-conjugated secondary antibodies (1:200), followed by counterstaining with a DNA-binding dye PI or DAPI (1 µg/mL in PBS) for 10 minutes. Fluorescence images were examined and photographed with the laser scanning confocal microscopy (Leica, Solms, Germany). About 8 vision fields were photographed randomly for each section. Each staining experiment was repeated at least 3–5 times.
Cell culture and treatment
Human gastric cancer cell lines MKN-45 and MGC-803 were cultured in an incubator at 37℃ in RPMI-1640 supplemented with 10% FBS with an atmosphere of 5% CO2. Prior to isoprenaline stimulation, cultures were incubated overnight in serum-free medium supplemented with 10 mM HEPES, 0.1% bovine serum albumin (pH 7.4). Cultures were then treated with 0, 5, or 10 uM isoprenaline for indicated time periods.
Quantitative real-time PCR analysis
Sixty-five gastric cancer and 42 normal gastric tissue samples were collected from the General Hospital of PLA with informed consent of patients and the institutional approval. The real-time polymerase chain reaction (PCR) was performed using the Mx3005P system (Stratagene) with a 10 µL of TaqMan Gene Expression Master Mix, 20 µL of reaction volume containing 5 µL of cDNA, 1 µL of TaqMan Gene Expression Assay primers and probes for periostin and glyceraldehyde-3-phosphate dehydrogenase (GAPDH). Primers for the genes of interest were: periostin193 bp (5'-GCCATCACATC GGACATA-3' and 5'-CTCCCATA ATAGACTCAGAACA-3'), and GAPDH 266 bp (5'-AGAA GGCTGGGGCTCATTTG-3' and 5'-AGGGGCCATCCACAGTCTTC-3'). The results were analyzed using the comparative threshold cycle (CT) method and normalized by a housekeeping gene GAPDH.
Western blotting analysis
Total proteins from cells were extracted with a ice-cold lysis buffer. The concentration of proteins in the supernatant was analysed by the bicinchoninic acid method. Protein samples were separated by SDS-PAGE and transferred to nitrocellulose membranes (Amersham Biosciences, Piscataway, NJ, USA). Membranes were blocked with 5% slim milk in TBST at room temperature for 1 hour. After blocking, blots were probed overnight at 4℃ with a primary antibody diluted in blocking buffer (anti-periostin antibody 1:1000, anti-β-actin antibody 1:1000). The membranes were incubated with a horseradish peroxidase conjugated goat anti-rabbit/mouse secondary antibody (1:1000) for 2 hours at room temperature after washing three times with TBST. The membranes were developed using enhanced chemiluminescence substrate (Pierce) to transfer to film (Millipore).
RESULTS
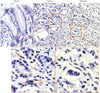
Periostin is expressed in the stroma of the primary tumors and metastases, but not expressed in normal gastric tissue
Periostin has been characterized as a component of the tumor microenvironment, or metastatic niche, capable of promoting cancer development and progression. However, expression of periostin in gastric cancer has not been sufficiently studied. In this study, we first performed in situ hybridization in peri-cancerous, cancerous, and metastatic gastric cancer tissue samples to examine the expression patterns of periostin at the transcription level. In normal gastric tissues, periostin expression was not detected in gastric epithelial cells or the stroma of gastric tissues (Fig. 1A). In cancerous tissues, however, periostin was upregulated and present in the stroma of the primary tumors, but not in gastric epithelial cells (Fig. 1B). In metastatic gastric cancer tissues, periostin was also detected in lymphatic metastasis sections (Fig. 1C).
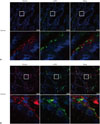
Periostin is mainly located in pericryptal fibroblasts but not in tumor cells
Stromal cells in gastric cancer produce periostin. Cancer-associated fibroblasts (CAFs), which interact with cancer cells to promote cancer progression, are identifiable by the expression of the myofibroblast marker α-SMA. Whether periostin in the cancer stroma is secreted by cancer cells or CAFs has not yet been determined in gastric cancer. We, therefore, collected gastric cancer tissue samples and used periostin combined with the fibroblast-associated protein α-SMA to label cancerous cells that co-expressed periostin and the fibroblast marker to evaluate the location of periostin in gastric cancer tissues. We first found that periostin expression correlated to the expression of α-SMA in gastric cancer tissues. Periostin was located mainly in the pericryptal fibroblasts (Fig. 2). In cancerous tissue where periostin was expressed at low levels, the expression of α-SMA was strongly correlated with the distribution patterns of periostin (Fig. 2A). In cancerous tissues where periostin showed high expression, α-SMA was also expressed at high levels in the membrane of cancerous gastric epithelial cells (Fig. 2B).
The distribution patterns of periostin increased as clinical staging of the tumor progressed
CAFs are the primary source of periostin, which facilitates tumor cell invasion by establishing a neoplastic niche in gastric cancers. Thus, we hypothesized that the niche increased its scope while the clinical staging of the tumor progressed. Given the active roles of periostin in regulating gastric cancer progression and metastasis, we then examined whether the distribution pattern of periostin was related to the clinical staging of tumors. Our results indicated that distribution patterns of periostin show a much broader distribution while the clinical staging of the tumor progressed (Fig. 3). Periostin is distributed at low levels in early gastric cancer (Fig. 3A). In intermediate gastric carcinoma, periostin shows a much broader distribution than in early gastric cancer (Fig. 3B). The distribution of periostin was the highest in late gastric cancer (Fig. 3C). These data suggest that the study of periostin may be a promising future research on the inhibition of gastric cancer progression and metastasis.
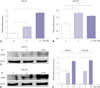
Isoprenaline upregulates expression levels of periostin in gastric cancer cells
An emerging role for stress-related hormones in regulating cancer progression has been recognized. However, whether stress serves as a mechanism to promote gastric cancer development is not clear. To further investigate the relationship between periostin and stress-related hormones, we first performed real-time PCR to evaluate periostin expression after isoprenaline treatment. Thus, MKN-45 and MGC-803 cells were incubated with increasing doses of isoprenaline, and periostin was evaluated by real-time PCR. After incubation with 10 µM isoprenaline, periostin mRNA levels increased remarkably with a 4.9-fold enhancement in MKN-45 and 1.4-fold increase in MGC-803 gastric cancer cells compared with control groups (Fig. 4A and B). Next, MKN-45 and MGC-803 cells were stimulated with 0, 5, or 10 µM isoprenaline after serum starvation and periostin expression was evaluated by Western blot analysis (Fig. 4C and D). Isoprenaline stimulation of MGC-803 cells also upregulated periostin expression at both mRNA and protein levels in a time-dependent manner (Fig. 5). Collectively, these observations showed that isoprenaline is a positive regulator of periostin activity in gastric cancer cells.
DISCUSSION
Invasion and metastasis of tumor cells is the main cause of death in patients with gastric cancer.21 The metastatic niche plays important roles in cancer development and progression.22 Periostin, a major niche component, is functionally involved in multiple steps of cancer progression and participates in different signaling pathways, and thus is indispensable for gastric cancer progression and metastasis.23 In this study, we found that periostin is expressed in the stroma of the primary tumors and metastases, but not in normal gastric tissue. We also found that α-SMA expression was significantly correlated with periostin; periostin was located mostly in pericryptal fibroblasts, but not in tumor cells, and strongly correlated to the expression of α-SMA. Since periostin is believed to change quantitatively or qualitatively in normal, primary cancer, and metastatic niches, we then investigated whether the distribution patterns of periostin was related to the clinical staging of tumor by immunofluorescence, and found that the distribution pattern of periostin was broader as the clinical staging of the tumor progressed.
A growing list of molecular cues can initiate tumor development and progression.24 Recently, the emerging role of stress-related hormones in the regulation of cancer development and progression has been recognized.25 Epidemiological studies indicate that stress-related hormones might serve as risk factors for cancer development.26 Thus, we next examined the expression levels of periostin in gastric cancer cells at both mRNA and protein levels after treatment with different doses of stress-associated hormone isoprenaline, and surprisingly found that isoprenaline enhanced the expression of periostin in gastric cancer cell lines. Isoprenaline stimulation also upregulated periostin expression in a time-dependent manner in MGC-803 cells, indicating a direct connection between stress-related hormones and gastric cancer development and progression.
Our data together our data suggest that isoprenaline recognized and activated its target receptors, subsequently activating the downstream target periostin. The activation of periostin by isoprenaline functions as a regulator of subsequent cancer progression, and is associated with poor prognosis in human gastric carcinoma.23 Our findings provide a biochemical mechanism involved in psychosocial influences on gastric cancer pathogenesis, and suggest that pharmacological interventions targeting periostin or isoprenaline could potentially be used to ameliorate stress-associated influences on gastric cancer development and progression. As gastric cancer treatment develops towards a more patient-specific direction, consideration of the influence of psychosocial factors provides a novel perspective for new therapeutic targets.


 XML Download
XML Download