

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Neisseria gonorrhoeae (N. gonorrhoeae) is the causative agent of gonorrhea, a sexually transmitted disease, and is one of the most prevalent sexually-transmitted pathogens worldwide.1 The incidence of N. gonorrhoeae infection has decreased in many developed countries,2 although it is still prevalent in many developing counties,1 and has even recently increased in a few developed countries.3 According to the Centers for Disease Control and Prevention (CDC) in the United States, a total of 321849 cases of gonococcal infections were reported in 2011; however, the CDC estimates that the incidence could exceed 800000 cases annually in the United States alone.4 In Korea, the Health Insurance Review and Assessment Service reported that 40038 cases of gonococcal infections were charged for pay in 2012.5
N. gonorrhoeae is considered to be a pathogen, and can only be transmitted from human to human via various sexual practices.6 Thus, infection control is very important to prevent the dissemination of N. gonorrhoeae among the general population. However, most female and a few male patients who have gonorrhea do not exhibit any apparent symptoms,7 despite the clinical significance of this disease. The lack of obvious symptoms therefore adds a layer of difficulty to controlling its spread. Furthermore, the emergence and spread of resistance to various antimicrobial agents has created many difficulties in the management of gonococcal infections.89 Drug-resistant N. gonorrhoeae was classified as a microorganism with a threat level of "urgent" by the CDC in 2013.4 Thus, epidemiological understanding and surveillance programs may need to be implemented in order to prevent the dissemination of gonorrhea and overcome the problem of antimicrobial resistance in N. gonorrhoeae.10
Tetracycline is a historic drug and cannot be used any longer to treat gonorrhea in many countries due to the dissemination of tetracycline-resistant strains of N. gonorrhoeae, which have become very common over the past several decades, including in Korea.11 Since tetracycline is not used anymore, the detection of tetracycline resistance is clinically insignificant. However, surveillance of trends in high-level resistance to tetracycline caused by plasmid transfer can be helpful in understanding the epidemiological aspects of gonococcal infections and controlling these infections, because such monitoring enables the detection of clonal variations.12 In this study, we aimed to determine the incidence of high-level tetracyclineresistant N. gonorrhoeae (TRNG) isolates over many years and to investigate the characteristics and genetic epidemiology of TRNG in Korea.
MATERIALS AND METHODS
N. gonorrhoeae isolates
A total of 601 N. gonorrhoeae isolates were collected from symptomatic patients with urethritis and sex workers between 2004 and 2011 in Korea. The number of isolates collected each year ranged from 47 to 135. Most N. gonorrhoeae isolates were obtained from primary urological clinics, which were widely distributed across the country. Modified Thayer-Martin agar plates (Becton Dickinson, Cockeysville, MD, USA) were used for primary isolation, and conventional biochemical tests, including Gram stain and the Vitek NHI System (bioMerieux, Marcy l'Etoile, France), were used for species identification. Isolates were stored in 20 per cent skim milk (Difco, Detroit, MI, USA) at -70℃ until ready for analysis.
Antimicrobial susceptibility testing
High-level resistance to tetracycline was tested by the disk diffusion and agar dilution methods. The disk diffusion test, which is recommended by the Clinical and Laboratory Standards Institute (CLSI), was performed using a disk containing 30 µg of tetracycline (Becton Dickinson).1314 Fifty-eight isolates of TRNG and 304 isolates of non-TRNG were randomly selected to determine the minimum inhibitory concentrations (MICs) of various antimicrobial agents, including tetracycline, and to compare resistances. Resistance to penicillin G (Sigma Chemical, Saint Louis, MO, USA), ceftriaxone (Hanmi, Seoul, Korea), spectinomycin (Kuk Je, Seoul, Korea), cefixime (Dong-A, Seoul, Korea), tetracycline (Pfizer Korea, Seoul, Korea), and ciprofloxacin (Bayer Korea, Seoul, Korea) were tested by the CLSI agar dilution method,1315 using a GC II agar base supplemented with 1% IsoVitaleX (Becton Dickinson). Approximately 104 colony forming unit of each isolate were inoculated onto the agar plates with a Steer's replicator (Craft Machine, Chester, PA, USA); plates were then incubated in a 5% CO2 atmosphere at 35℃ for 24 h. The American Type Culture Collection (ATCC) 49226, World Health Organization (WHO) A 100A6, WHO B 100A7, WHO G 100I1, WHO J 100I4, WHO K 300J4, and WHO L 00G 1003 OKKA20 strains of N. gonorrhoeae were used as quality control strains.
Molecular epidemiological study
To determine the molecular epidemiological relatedness of the TRNG strains, N. gonorrhoeae multi-antigen sequence typing (NG-MAST) and pulsed-field gel electrophoresis (PFGE) were carried out for 58 and 36 isolates of TRNG, and NG-MAST for 156 isolates of non-TRNG was tested.12 NG-MAST was performed according to the guidelines outlined on the NG-MAST homepage (http://www.ng-mast.net), using primers for the porB and tbpB genes.16 Polymerase chain reaction (PCR) was carried out in a total volume of 20 µL, containing 1 µL of heatextracted template DNA, 10 pmol of each primer, and a PreMix (Bioneer, Daejeon, Korea) master mix containing 1 U of Taq DNA polymerase. Thermocycling conditions included 25 cycles of 95℃ for 30 seconds, either 58℃ for 30 seconds for porB or 69℃ for 30 seconds for tbpB, and 72℃ for 1 minute. Reactions also included an initial denaturation step at 95℃ for 4 minutes and a final extension step at 72℃ for 10 minutes, and were performed using a thermal cycler (Eppendorf, Hamburg, Germany). The resultant PCR products were purified using a DNA extraction kit (Qiagen, Hilden, Germany), and the nucleotide sequences of the PCR-generated amplicons were analyzed at a commercial laboratory (Macrogen, Seoul, Korea). The sequences of porB and tbpB were analyzed using the NG-MAST homepage to assign allele numbers and to determine the sequence type (ST) of each isolate. Two phylogenetic trees of por genes for TRNG and non-TRNG and tbpB genes for TRNG were generated with Molecular Evolutionary Genetics Analysis version 6 software (http://www.megasoftware.net) using the Neighbor-Joining method.
PFGE was performed for 36 isolates, which showed different NG-MAST STs. Briefly, one loopful of cells was suspended in 1 mL of saline ethylenediaminetetraacetic acid (EDTA) solution and used to prepare genomic DNA plugs, which were digested with NheI (Takara, Tokyo, Japan) for 18 hours at 35℃. Fragments were then resolved using switch times of 0.5 seconds (initial) and 54 s (final), and a running time of 20 h at 6 V/cm with a CHEF DR II instrument (Bio-Rad, Hercules, CA, USA). PFGE banding pattern similarities were determined using the Dice coefficients and the unweighted pair group method, using arithmetic averages obtained by the clustering method. Calculations were performed using Molecular Analyst Fingerprinting software (v. 1.12; Bio-Rad), and dendrogram was generated from these calculations.
RESULTS
Incidence of TRNG
A total of 92 isolates (15%) of TRNG were identified from 2004 to 2011. The proportion of TRNG was low in 2004 (2%, 2/91); however, it increased to 10% (5/48) in 2005. This incidence of TRNG continued to steadily increase, reaching 24% (16/66) in 2007. The range of the incidence of TRNG from 2008 to 2011 was 15–21% (Fig. 1).
Antimicrobial susceptibility
None of the non-TRNG isolates were susceptible to penicillin G; however, all of the non-TRNG isolates showed susceptibility to ceftriaxone, cefixime, and spectinomycin. The MIC50 and MIC90 of ceftriaxone and cefixime were 0.06 mg/L and 0.12 mg/L, respectively. The MIC50 and MIC90 of ciprofloxacin were 8 mg/L and 16 mg/L, and 15% of all isolates were susceptible to ciprofloxacin. Like the non-TRNG isolates, none of the TRNG isolates were susceptible to penicillin G; meanwhile, all TRNG isolates were susceptible to ceftriaxone, cefixime, and spectinomycin. However, the MIC50s and MIC90s of ceftriaxone and cefixime were very low (≤0.008 mg/L), unlike for the non-TRNG isolates. The rate of susceptibility to ciprofloxacin among the TRNG isolates was 61%, and the MIC50 and MIC90 of ciprofloxacin were ≤0.008 mg/L and 4 mg/L, respectively (Table 1).
From 2004 to 2011, no definite changes in antimicrobial susceptibility were noted in the non-TRNG isolates; however, the susceptibility to cephalosporins and the rate of susceptibility to ciprofloxacin deteriorated in the TRNG isolates. The MIC50 and MIC90 of ceftriaxone were ≤0.008 mg/L and ≤0.008 mg/L in 2004–2007, ≤0.008 mg/L and 0.06 mg/L in 2008–2009, and 0.03 mg/L and 0.06 mg/L in 2010–2011, respectively. The MIC50 of ciprofloxacin and rate of susceptibility were ≤0.008 mg/L and 93% in 2004–2007, 2 mg/L and 44% in 2008–2009, and 4 mg/L and 18% in 2010–2011, respectively (Table 1).
Molecular epidemiology
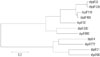
From 2004 to 2007, five different STs were found in 12 isolates; of these, six isolates belonged to ST1798 (50%) according to NG-MAST analysis, and nine isolates contained the tbpB 455 allele. All isolates collected in this period were highly susceptible to both ceftriaxone and cefixime, and all isolates except one showed susceptibility to ciprofloxacin. During 2008 and 2009, the STs of the isolates became more variable, with 13 different STs observed in 24 isolates; however, ST1798 remained the most prominent ST (7/24). Most of the N. gonorrhoeae isolates harboring the tbpB 455 allele showed susceptibility to ciprofloxacin, but other isolates that harbored the tbpB 21, 33, 60, 66, 110, or 272 allele were either intermediately or completely resistant. During 2010 and 2011, 15 different STs were noted; moreover, no two isolates shared the same ST. The most common tbpB allele was 21 (7/15), and only three isolates harbored the tbpB 455 allele, even though this allele had been highly prevalent during previous periods. Most of the isolates were resistant to ciprofloxacin, except the ST7934 and ST7924 isolates (Table 2). Dendrogram analysis revealed that most isolates were genetically unrelated, and suggested the sporadic emergence of various STs from 2007 to 2011. Some isolates harboring the same tbpB allele belonged to various STs, which were located distantly from one another on the dendrogram of TRNG (data not shown). In the phylogenetic tree, which was composed for the por gene for TRNG and non-TRNG isolates in 2006, three TRNG isolates had por 1146, while another exhibited por 2392; both alleles were genetically related to each other. However, the por gene of TRNG varied during 2010 and 2011, and the location of por gene alleles were dispersed throughout the phylogenetic tree (Fig. 2). In the phylogenetic tree of tbpB genes, which was composed for the 11 tbpB alleles identified from 2004 to 2011, two clusters were noted. The large cluster consisted of tbpB 33, 60, 110, 129, 330, 455, and 860, whereas the smaller cluster comprised tbpB 4, 21, 66, and 272 (Fig. 3).
DISCUSSION
The dissemination of multidrug-resistant strains of N. gonorrhoeae and the recent emergence of resistance to extendedspectrum cephalosporins, including ceftriaxone, pose major problems for the management of gonorrhea.171819 A particular difficulty in achieving effective antimicrobial treatment is that it requires active infection control, based on precise epidemiological information, to overcome gonorrheal infections. Tetracycline resistance can be caused by chromosomal mutations in several genes, such as mtrR, or by the transfer of plasmids harboring the tetM gene.2021
In Korea, tetracycline resistance has been widely disseminated since the late 1960s, and almost all isolates have been tetracycline-resistant for several decades.112223 However, in spite of the prevalence of tetracycline resistance, the incidence of TRNG remained relatively low until the early 2000s, ranging from 0–1% in the late 1990's and 1–4% in the early 2000's.1222 In 2004, the incidence of TRNG was still low (2%) in Korea; however, high-level tetracycline resistance increased to 10% in 2005 and again up to 24% in 2007. The reasons underlying this increase in TRNG isolates are not yet clear. However, hypotheses can be generated by taking into account clonal spread, horizontal spread, and foreign import. First, clonal spread may have contributed to the rise of TRNG in Korea. In antimicrobial susceptibility comparisons, TRNG isolates showed differences in their cephalosporin and ciprofloxacin susceptibilities, compared with non-TRNG isolates. The MIC50s and MIC90s of ceftriaxone and cefixime for TRNG isolates were eight times lower than for non-TRNG isolates; furthermore, the rate of ciprofloxacin susceptibility was 61% in TRNG isolates, but only 15% in non-TRNG isolates. This difference can be interpreted in the context of clonal differences and expansion of TRNG clones; moreover, clonal spread of TRNG isolates has been observed in the other countries including UK and Argentina.2425 The importance of clonal dissemination is also supported by the observation that most of the TRNG isolates obtained from 2004 to 2007 were of the ST1798 subtype and harbored the tbpB 455 allele (ST1798, ST6327, and ST7927); moreover, these isolates were placed into one common phylogenic cluster, shared with the tbpB 33 (ST7894) and tbpB 860 (ST3945) alleles.
However, the susceptibilities of TRNG isolates to ceftriaxone and ciprofloxacin were not consistent, and decreased over time. From 2004 to 2007, the MIC50s and MIC90s of TRNG isolates to ceftriaxone and cefixime were lower than 0.008 mg/L. From 2008 to 2009, the MIC90s of ceftriaxone and cefixime increased to 0.06 mg/L; moreover, the MIC50s of ceftriaxone and cefixime also increased to 0.03 mg/L and 0.06 mg/L, respectively. The MIC50 of ciprofloxacin and the rate of resistance to ciprofloxacin also increased over time. These changes in the antimicrobial susceptibilities of TRNG isolates were also reported in India by Bala, et al.26 Such changes can result from the horizontal spread of a plasmid bearing the tetM gene to resistant non-TRNG isolates, because plasmids can move between isolates.272829 This phenomenon may provide a second explanation for the increase in TRNG isolates. However, considering the fact that the selective pressure for tetracycline was very low in Korea since the 1990s,22 horizontal transfer seems a less likely explanation than the other alternatives.
Notwithstanding, many isolates of TRNG were located at a great distance from one another in the dendrogram and phylogenetic tree in the period of increasing prevalence of TRNG. In 2006, three of four TRNG isolates belonged to ST1798 harboring por 1146 allele, while the remaining isolate exhibited por 2392, revealing a genetic relationship indicative of clonal expansion of TRNG. However, this clonal relationship was not found during 2010 and 2011. TRNG collected during this period seemed to appear sporadically, and their appearance is not easily explained by clonal spread. During this period, half of the por alleles (8/16) were located outside of the main cluster that consisted of por 23, 2053, 2874, 4270, 4801, 4917, 6288, and 6297, which may indicate emergence of divergent TRNG strains. The emergence of these divergent isolates can suppose that the increase of TRNG prevalence can result from foreign import when we take into consideration that consistent collection strategies have been sustained since 2004. This hypothesis also can be supported by a change in the tbpB allele throughout the late 2000s in Korea. From 2004 to 2007, the majority of all TRNG isolates in Korea belonged to STs bearing the tbpB 455 allele (ST1798, ST6327, and ST7927); however, STs harboring the tbpB 21 allele (ST7689 and ST7928) emerged in 2008, making this allele the most prominent allele during 2010 and 2011. Strains harboring the tbpB 455 or tbpB 21 allele showed many differences in their individual allele sequences, and this can suggest the emergence of new STs resulting from foreign import.30 In the Asia-Pacific region, the proportion of TRNG isolates varies according to country.3132 In 2004, 72% of all N. gonorrhoeae isolates in Singapore were TRNG, 34% in China, 20% in Vietnam and 8% in the Philippines. Japan exhibited a low incidence (2%) of TRNG, similar to that of Korea.31 In 2010, the incidence of TRNG isolates in Western-Pacific countries ranged from moderate to high, with incidences of 71–100% in Brunei; 35–75% in Mongolia, China, Hong Kong, Singapore, and Vietnam; and 10–34% in Australia, India, the Philippines, and Thailand.32 It is difficult to thoroughly assess the extent of foreign import of some pathogens; however, the recent worldwide spread of cefixime-resistant strains of N. gonorrhoeae33 and genetic relatedness of ceftriaxone-resistant strains isolated from France and Spain may be a convincing examples of foreign import.34 In our study, the emergence of new STs, the changes in tbpB allele distribution, the dispersed distribution of isolates sharing the same tpbB allele in the dendrogram, and the dispersed distribution of isolates of various STs according to year in the dendrogram appear to support the idea of foreign import. This import of TRNG strains from other countries can be a source of another severe antimicrobial resistance, including ceftriaxone resistance, and may require additional programs to prevent it. However, further investigations employing additional approaches, such as multilocus sequence typing, should be performed to test this hypothesis.
In conclusion, the increase in the incidence of TRNG strains until 2007 appears to be due, at least in part, to clonal spread. However, we propose that the emergence of various STs since 2008 is due to foreign import and/or clonal evolution. Further investigations are required to fully determine the relevant changes in TRNG epidemiology.




 XML Download
XML Download