

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Mammalian cells usually uptake macromolecules from the extracellular microenvironment through their receptor(s) or by penetration.1 Professional phagocytes such as macrophages (Mϕ) are not exceptional. However, Mϕ have another unique uptake process called phagocytosis, in which Mϕ actively uptake not only macromolecules but also large particles such as bacterial pathogens.1
Mϕ recognize and engulf invading bacteria, and specific vacuoles called phagosomes are then formatted therein.2 The phagosomes mature into phagolysosomes, in which bacterial pathogens encounter various antimicrobial agents such as lysozymes by which bacterial pathogens are digested.3 Reactive oxygen intermediates and nitrogen oxide synthesized in the cytosol participate in the killing of bacterial pathogens by penetrating into phagolysosomes.4
Since Mϕ are particularly important for protection against bacterial infection, analysis of the phagocytic and bactericidal activities of Mϕ is essential for the determination of their functional activities. Bacterial pathogens are categorized into at least two groups on the basis of their kinetics in Mϕ:5 extracellular bacteria and intracellular bacteria. Extracellular bacteria are easily killed by Mϕ, whereas intracellular bacteria show resistance to digestion by Mϕ.5 However, the majority of intracellular bacteria are also killed by Mϕ, although this depends on the activation status of Mϕ.678910
Several methods are employed to determine the phagocytic and bactericidal activities of Mϕ. Yet, in most cases, expensive materials and equipment are usually required, and the methods are rather complicated. Therefore, this review focuses on a simple, reproducible, inexpensive, yet old-fashioned method for determining the phagocytic and bactericidal activities of Mϕ.
WHAT IS AN ANTIBIOTIC PROTECTION ASSAY?
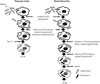
An antibiotic protection assay is traditionally employed to determine the phagocytic and bactericidal activities of Mϕ.678910111213 This assay system is based on counting colony-forming units (CFU) in Mϕ after phagocytosis (Fig. 1). After incubating Mϕ with bacteria for a short period of time, the bacteria are engulfed by Mϕ. The phagocytic activity of Mϕ can be determined by counting the CFU in Mϕ at this time point [CFU (P)]. After further incubation for a short period of time, the engulfed bacteria are killed by Mϕ. The number of viable bacteria in Mϕ can be counted at this time point [CFU (B)]. The bactericidal activity of Mϕ can thus be calculated by comparing CFU (P) with CFU (B). Thus, both the phagocytic and bactericidal activities of Mϕ can easily be determined. It is important that bacteria not engulfed by Mϕ must be killed to avoid bacterial growth outside the Mϕ. As antibiotics are essential for killing bacteria that are not engulfed by Mϕ, this assay is known as an antibiotic protection assay.
ANTIBIOTICS RECOMMENDED FOR AN ANTIBIOTIC PROTECTION ASSAY
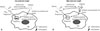
In an antibiotic protection assay, selection of an antibiotic that can effectively kill extracellular (i.e., bacteria not engulfed by Mϕ) yet not intracellular (i.e., bacteria engulfed by Mϕ) bacteria is quite important. As the sensitivity against antibiotics differs for each bacterium, an antibiotic that can effectively kill bacteria must be employed. Antibiotics with low molecular weight pass through plasma membrane of Mϕ. Therefore, the bacteria engulfed by Mϕ are killed by this type of antibiotic (Fig. 2A). In contrast, antibiotics with high molecular weight are unable to penetrate into cytosol. Therefore, the bacteria engulfed by Mϕ are not killed by this type of antibiotic (Fig. 2B). Thus, an antibiotic with a high molecular weight must be used in an antibiotic protection assay.
GENTAMICIN (GM) IS WIDELY USED FOR DETERMINING PHAGOCYTIC AND BACTERICIDAL ACTIVITIES OF Mϕ IN AN ANTIBIOTIC PROTECTION ASSAY
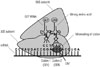
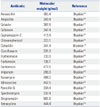
Numerous antibiotics with high molecular weight have been identified previously.14 Among these, gentamicin (GM) is widely employed for an antibiotic protection assay. The reasons are as follows: 1) GM is an aminoglycoside antibiotic that binds to the bacterial ribosome 30S subunit and induces the misreading of a wide range of RNAs (Fig. 3).15 2) GM has a broad-spectrum (Table 1).1617181920 3) GM is considered to be unable to penetrate into the cytosol of Mϕ due to its high molecular weight (Table 2).14 It is generally accepted that molecules with a molecular weight of less than 400 g/mol are able to pass through the plasma membrane.21 Therefore, antibiotics with a molecular weight of more than 400 g/mol are recommended for determining the phagocytic and bactericidal activities of Mϕ. There are several antibiotics with a higher molecular weight than that of GM (Table 2). However, these antibiotics are not recommended by several reasons. For example, the molecular weight of erythromycin is markedly higher than that of GM, yet erythromycin expresses bacteriostatic, but not bactericidal, activity.14 Similarly, the molecular weights of kanamycin and streptomycin are higher than those of GM (Table 2).14 However, Pseudomonas spp. show resistance to these antibiotics.2223 It is needless to say that antibiotics other than GM with high molecular weight can nevertheless be employed for determining the phagocytic and bactericidal activities of Mϕ against particular bacterial pathogens. However, GM is recommended for determining these activities, as this antibiotic has a broad spectrum and kills extracellular, but not intracellular, bacteria. Therefore, we focus on an antibiotic protection assay using GM (GM protection assay) in the following section.
EXPERIMENTAL PROCEDURE FOR DETERMINING PHAGOCYTIC ACTIVITY OF Mϕ USING A GM PROTECTION ASSAY
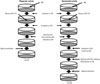
An experimental procedure for determining the phagocytic activity of Mϕ using a GM protection assay is shown in Fig. 4. Mϕ are incubated in RPMI 1640 containing 10% fetal calf serum [designated as complete medium (CM)] for 120 min to adhere to the bottom of tissue culture plates. Cells are incubated with bacteria for a given length of time to ingest bacteria. Subsequently, cells are washed three times with CM containing the optimal concentration of GM to remove non-ingested bacteria. Note that pre-warmed, but not cold, CM should be used throughout the experiment so as not to detach the Mϕ from the bottom of tissue culture plates. After washing with CM, a portion of cells is treated with saponin, which is plated on agar plates after sonication, and the CFUs are determined. Phagocytic activity is calculated as follows: {number of viable bacteria ingested by Mϕ [CFU (P)]/total number of viable bacteria incubated with Mϕ [CFU (T)]}×100 (%). Thus, the percentage of bacteria engulfed by Mϕ (phagocytic activity) can be quantitated.
EXPERIMENTAL PROCEDURE FOR DETERMINING BACTERICIDAL ACTIVITY OF Mϕ USING A GM PROTECTION ASSAY
An experimental procedure for determining the bactericidal activity of Mϕ using a GM protection assay is also shown in Fig. 4. Mϕ incubated in CM for 120 min are incubated with bacteria and then washed three times with CM containing the optimal concentration of GM to remove non-ingested bacteria. Subsequently, cells are further incubated in CM containing the optimal concentration of GM for a given length of time to kill non-ingested bacteria followed by CM. Cells are then treated with saponin, plated on agar plates after sonication, and the CFUs are determined. Bactericidal activity is calculated as follows: 100-{number of remaining viable bacteria in Mϕ [CFU (B)]/CFU (P)×100} (%). Thus, the percentage of bacteria killed by Mϕ (bactericidal activity) can be quantitated.
AN ANTIBIOTIC PROTECTION ASSAY FOR BACTERIA SHOWING RESISTANCE TO GM
As described above, GM is widely used to determine the phagocytic and bactericidal activities of Mϕ against various bacteria. However, certain bacteria show resistance to GM (Table 3).15,17 Hence, GM cannot be used to determine the phagocytic and bactericidal activities of Mϕ against these bacteria. In such cases, an antibiotic other than GM with a high molecular weight must be used.
PRELIMINARY EXPERIMENTS THAT SHOULD BE PERFORMED BEFORE A GM PROTECTION ASSAY
Although the GM protection assay is quite simple, preliminary experiments must be performed. One of the most important points is to determine the optimal conditions (i.e., concentration and length of effectiveness) of GM. Although GM has been considered to be unable to kill bacteria in Mϕ,1213242526 several studies have reported that GM, even in high concentrations, kills bacteria in Mϕ.2728 Therefore, the optimal concentration and length of effectiveness for GM should be determined with care. After determining the minimal inhibitory concentration (MIC), the optimal concentration of GM is then determined. Specifically, Mϕ infected with bacteria are incubated with different concentrations of GM (higher than MIC; e.g., 2.5-fold MIC), and verification that GM does not penetrate into Mϕ should be performed. In addition, further verification as to whether Mϕ are damaged by the concentration of GM should also be performed, as Mϕ are commonly destroyed by high concentrations (e.g., 100 µg/mL) of GM (Fig. 5).
Saponin facilitates the destruction of plasma and phagosomal membranes of Mϕ by interacting with cholesterols, which are abundant in their plasma membranes (Fig. 6).29 Therefore, before counting CFU in Mϕ, Mϕ must be treated with saponin. We can confirm that Mϕ are completely destroyed by 0.5% saponin.
In order to determine the phagocytic and bactericidal activities of Mϕ, the CFUs in Mϕ are counted at different time points after infection. In most cases, the number of viable bacteria engulfed by Mϕ is highest at 45-60 min after incubation with bacteria, and the bacteria are usually killed within 90-120 min after being engulfed by Mϕ. It is needless to say that there are some exceptions; for example, Mycobacterium spp. are not killed within 120 min after being engulfed by Mϕ.3031
SEVERAL METHODS RECENTLY EMPLOYED FOR DETERMINING PHAGOCYTIC AND BACTERICIDAL ACTIVITIES OF Mϕ
Bacteria engulfed by Mϕ can be detected using a light microscope, a fluorescence microscope, or a flow cytometer.3233343536 However, it is difficult to distinguish bacteria engulfed by Mϕ from those merely attached to the plasma membranes of Mϕ by these methods. The problem can be solved by using ethidium bromide.32333536 Although the phagocytic activity of Mϕ can be determined using these methods, another experiment must be performed to determine the bactericidal activity of Mϕ using MTT and an absorption spectrometer in each case.3336373839 Thus, it is possible to determine the phagocytic and bactericidal activities of Mϕ using these methods. However, expensive materials and equipment are required and the methods are rather complicated. Moreover, it is impossible to determine both the phagocytic and bactericidal activities of Mϕ simultaneously using these methods.
CONCLUSION
This review describes a simple, reproducible, inexpensive, yet old-fashioned method for determining the phagocytic and bactericidal activities of Mϕ. Although the phagocytic and bactericidal activities of Mϕ against various bacterial pathogens can be determined by using methods that have been recently employed, it is impossible to determine both activities simultaneously. In addition, these methods require expensive materials and equipment and complicated methods. In contrast, both the phagocytic and bactericidal activities of Mϕ can be determined simultaneously by using an antibiotic protection assay for which expensive materials and equipment are not required. We therefore recommend investigators to reevaluate the antibiotic protection assay.



 XML Download
XML Download