

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Helicobacter pylori (H. pylori) infection mediates gastritis and gastric adenocarcinoma.1 Interleukin-8 (IL-8) contributes to gastric inflammation.2 IL-8 levels are found to be elevated in gastric mucosal tissues of the patients infected with Helicobacter pylori2 and H. pylori-infected gastric epithelial cells.3,4 Reactive oxygen species (ROS) mediate the expression of IL-8 by activating oxidant-sensitive transcription factors, such as nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), activator protein-1 (AP-1), janus kinase/signal transducers, and activators of transcription (Jak/Stat).345
H. pylori exhibits chemotactic activity by inducing neutrophil activation, and these activated neutrophils induce ROS production.67 It was recently reported that ROS is involved in Jak/Stat signal molecules in inflammatory signaling pathway of non-phagocytic cells, as well as phagocytic cells. Jak/Stat signaling mediates activation of cytokine signaling.89
There are three subfamilies of mitogen-activated protein kinases (MAPKs); extracellular signal-regulated kinases (ERKs), c-Jun NH2-terminal protein kinases (JNKs), and p38 MAPK. The cytotoxin-associated gene (cagA) pathogenicity island of H. pylori is involved in NF-κB and MAPK activation in gastric epithelial cells.3 Transcription of IL-8 gene requires NF-κB activation and NF-κB is indispensable for the enhanced IL-8 mRNA transcription in H. pylori-infected gastric epithelial cells.347
α-lipoic acid (α-LA) is supplied from diets such as spinach and broccoli and from a supplement. α-LA and its active reduced counterpart dihydrolipoic acid (DHLA) reduce oxidative stress by chelating transition metals, recycling endogenous antioxidants, and scavenging ROS.1011 α-LA showed beneficial effect on treating ROS-mediated diseases including diabetes, atherosclerosis, and hypertension.12131415
Therefore, we investigated whether α-LA reduces levels of ROS produced in H. pylori-infected gastric epithelial cells, thereby suppressing the activation of inflammatory signaling molecules, such as MAPK (ERK1/2, JNK1/2, p38), Jak/Stat (Jak1, Jak2, Stat3), transcription factor NF-κB, and IL-8 expression in H. pylori-infected gastric epithelial cells.
A human gastric epithelial cell line AGS (gastric adenocarcinoma, ATCC CRL 1739) was purchased from the American Type Culture Collection (Manassas, VA, USA) and cultured as previously described.3 H. pylori strain in a Korean isolate (HP99; cagA+, vacA s1b, m2, iceA genotype) was inoculated onto chocolate agar plates at 37℃ under microaerophilic conditions using GasPak™ EZ Gas Generating Pouch Systems (BD Biosciences, San Jose, CA, USA).3 Prior to infection, H. pylori were harvested, and then resuspended in antibiotic-free cell culture medium. H. pylori was added to cultured cells at a bacterium/cell ratio 300:1.
For time-course experiment for IL-8 levels, cells were infected with H. pylori for several time points. α-LA was purchased from Sigma-Aldrich (St. Louis, MO, USA) and dissolved in ethanol. The cells were pretreated with α-LA (final concentrations of 10 and 20 µM) for 2 h and then infected with H. pylori for 30 min (for ROS levels, NF-κB, p-IκBα, IkBα, MAPK, Jak/Stat), 3 h (for IL-8 mRNA) or 12 h (for IL-8 protein levels). None and control cells without α-LA received ethanol instead of α-LA. The time points for determining ROS, NF-κB, p-IκBα, IκBα, MAPK, and Jak/Stat, as well as 2 h-pretreatment of α-LA, were adapted from our previous studies.161718
IL-8 levels in the medium were determined by using enzyme linked immunosorbent assay (ELISA) kits (Biosource International, Inc., San Diego, CA, USA) following the manufacturer's instructions. For real-time PCR analysis, total RNA in cells were isolated and converted into cDNA by reverse transcription process using a random hexamer and virus reverse transcriptase (Promega, Madison, WI, USA). Sequences of IL-8 primers and β-actin were adapted from our previous study.19 cDNA was added in a SYBR Green Realtime PCR Master Mix (TOYOBO Co., Osaka, Japan) containing 10 pg/mL of forward and reverse primers for IL-8. cDNA was amplified by 40 cycles, denaturation at 95℃ for 15 sec, annealing at 60℃ for 5 sec, and extension at 72℃ for 30 sec. β-actin gene was amplified in the same reaction to serve as the reference gene.
ROS levels were determined using 2',7'-dichlorodihydrofluorescein diacetate (Invitrogen, Carlsbad, CA, USA) as previously described.20 The amount of ROS trapped in the cells was expressed as the relative increase over the ROS level in cells cultured in the absence of H. pylori, which was set at 100.
NF-κB-DNA binding activity was determined by electrophoretic mobility shift assay (EMSA) as previously described.4
For Western blot analysis, proteins in whole cell extracts were subjected to 6% sodium dodecyl sulfate (SDS)-polyacrylamide gel electrophoresis, and transferred to nitrocellulose membranes which was blocked using 3-5% nonfat dry milk in Tris-buffered saline and 0.2% Tween 20 (TBS-T) for 2 h at room temperature. The membranes were incubated with antibodies for total and phospho-specific forms of ERK1/2, JNK1/2, p38, Stat3, Jak1, and Jak2 (Santa Cruz Biotechnology, Santa Cruz, CA, USA) diluted in TBS-T containing 3% dry milk at 4℃ for 16 h. After washing with TBS-T, primary antibodies were detected using horseradish peroxidase-conjugated secondary antibodies (anti-goat or anti-rabbit), respectively, and visualized by the enhanced chemiluminescence (ECL) detection system (Santa Cruz Biotechnology) according to the manufacturer's instruction.
The statistical differences were determined using one-way ANOVA and Newman-Keul's test. All values are expressed as mean±standard error of means (SEM) of four different experiments. A value of p<0.05 was considered statistically significant.
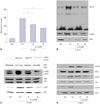
As shown in Fig. 1A and B, H. pylori infection induced mRNA expression of IL-8 time-dependently. IL-8 levels in the medium were also increased by H. pylori infection. H. pylori-induced increases in IL-8 mRNA levels were maximized at 3 h, while IL-8 protein levels in the medium stably increased from 8 h-culture. Therefore, to examine the effect of α-LA on H. pylori-induced expression of IL-8, AGS cells were pretreated with α-LA and cultured in the presence of H. pylori for 3 h (to assess mRNA levels) and 8 h (to assess protein levels in the medium) (Fig. 1C and D). α-LA showed inhibitory effect on H. pylori-induced IL-8 expression at both mRNA and protein levels in a dose-dependent manner.
To investigate whether α-LA inhibits H. pylori-induced increases in ROS levels and activation of NF-κB, intracellular ROS levels, NF-κB DNA binding activity, and levels of total and phospho-specific forms of IκBα were determined in the cells infected with H. pylori treated with or without α-LA. α-LA suppressed H. pylori-induced increases in ROS levels and NF-κB activation, accompanied with inhibition of phosphorylation of IκBα and a decrease in the total forms of IκBα (Fig. 2A and B). As shown in Fig. 2C, α-LA suppressed H. pylori-induced phosphorylation of ERK1/2, JNK1/2, and p38, while total forms of ERK1/2, JNK1/2, and p38 were not affected by H. pylori infection or α-LA treatment.
We investigated whether Jak-Stat signaling is involved in H. pylori-induced inflammation. As shown in Fig. 2D, H. pylori infection induced phosphorylation of Stat3, accompanied with phosphorylation of Jak1 and Jak2 in AGS cells, which was inhibited by α-LA dose-dependently. Total forms of Stat3, Jak1, and Jak2 were not changed by H. pylori infection or α-LA treatment. The results suggest that MAPK and Jak1/2-Stat3 pathways mediate H. pylori-induced IL-8 expression, which is inhibited by α-LA.
In the present study, we found that H. pylori, Korean isolate (HP99), induces IL-8 expression and activation of MAPK, Jak/Stat, and NF-κB, which were inhibited by α-LA. Since ROS mediates activation of MAPK, Jak/Stat, and NF-κB cells,1718 the inhibitory effect of α-LA on ROS production may suppress H. pylori-induced signaling for IL-8 expression in AGS cells. Several studies have reported that MAPK inhibitors, U0126 (an ERK inhibitor), and SB203580 (a p38 inhibitor) suppressed NF-κB activation in H. pylori-infected AGS cells.3 These results indicate that NF-κB activation acts as a downstream of ERK and/or p38 signaling in H. pylori-infected AGS cells. Therefore, α-LA may inhibit H. pylori-induced IL-8 expression through suppression of MAPK-mediated NF-κB activation in AGS cells.
Additionally, we found that α-LA inhibits H. pylori-induced Stat3 activation in AGS cells. Jak-Stat signaling is responsible for various cellular responses to cytokines, growth factors, and hormones.21 Bronte-Tinkew, et al.22 demonstrated that H. pylori activates Stat3 in gastric epithelial cells. Inhibition of Jak/Stat activation with chemical inhibitors suppresses phosphorylation of ERK, indicating that ERK/NF-κB signaling acts as a downstream of Jak2 activation.23 For phosphorylation of Stat3, activation of Jak is required. Therefore, Jak may be phosphorylated prior to activation of Stat3 by infection of H. pylori. In the present study, both Jak1 and 2 were phosphorylated along with phosphorylation of Stat3 in AGS cells. Since H. pylori induces actvation of MAPK and Jak/Stat at 30 min and NF-κB activation at 1 h-culture, Jak/Stat and MAPK may be upstream signaling of NF-κB in HP99-infected AGS cells. Since α-LA reduces ROS levels and activation of MAPK, Jak1/2-Stat3, and NF-κB, and thus, IL-8 expression, α-LA may have a therapeutic potential for H. pylori infection-associated inflammation.

 XML Download
XML Download