

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Photodynamic therapy (PDT) is a therapeutic modality using light and light-sensitive chemicals (photosensitizers). Its application has been successful in both neoplastic and non-neoplastic diseases.1 Recently, this therapy has been widely applied in various tissues and organs, including the retina, esophagus, stomach, skin, and colon. Krosl, et al.2 reported that the antitumor effect of PDT was improved by the administration of granulocyte-macrophage colony-stimulating factor (GM-CSF). In addition, certain gene transfections, such as Bcl-2 and IL-6, were reported to increase cellular sensitivity to PDT.34
Peroxisomal proliferator-activated receptor gamma (PPARγ) is a member of the nuclear receptor superfamily and plays a role in global cellular functions, including lipid and glucose metabolism, insulin sensitivity, adipocyte differentiation, proliferation, and apoptosis.5 Similar to the binding of ligands, PPARγ heterodimerizes with retinoid X receptor alpha and undergoes a conformational change that leads to the binding of a specific DNA sequence (peroxisomal proliferator response element).6 Recent reports indicate that, in addition to adipocytes, PPARγ is expressed at a significant level in various kinds of tumors, such as breast adenocarcinoma,789 colon adencarcinoma,10 liposarcoma,11 lung adenocarcinoma,12 and neuroblastoma.13 Many reports have demonstrated that activation of PPARγ by its specific ligand causes cancer cells to undergo apoptosis. It was also reported that PPARγ ligands induce growth retardation14 and apoptotic or non-apoptotic cancer cell death.15161718 However, the molecular mechanisms of PPARγ-induced apoptosis are currently under investigation.
Due to the limitation of the tolerable dose of PPARγ ligand in vivo, trials have been required to use a combination therapy with another cytotoxic drug. For example, the combination of PPARγ ligand with TNF-related apoptosis-inducing ligand (TRAIL) demonstrated an improved killing effect on cancer cells.1920 With this background, we asked whether treatment using troglitazone improves PDT tumor responsiveness and addressed this question by analyzing cell proliferation and apoptosis among DLD-1 colon cancer cells. Our results indicate enhanced tumoricidal activity when PDT is combined with the PPARγ ligand troglitazone.
MATERIALS AND METHODS
Cell lines and reagents
DLD-1, HCT-15, and HT-29 cells (human epithelial colon cancer cell lines, ATCC CCL 221) were cultured in RPMI 1640 medium supplemented with 10% fetal bovine serum (Gibco BRL, Grand Island, NY, USA). Pyropheophorbide-a methyl ester (PPME) was solubilized in N, N-dimethyl formamide (DMF). Troglitazone was provided from SanKyo (Tokyo, Japan) and adjusted to 10 mM in 19% (w/v) bovine serum albumin (BSA) and 5% (v/v) dimethyl sulfoxide (DMSO; Sigma, St. Louis, MO, USA) and added to the cell culture in a final concentration of 1 to 100 µM. Added to the control cultures was the same amount of DMSO with BSA, which was less than 0.2% (v/v) of the culture medium.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNA was prepared using an RNeasy Kit (Qiagen Inc., Chatsworth, CA, USA). cDNA was synthesized from 5 µg of total RNA using 2 µg random hexamer (Amersham Pharmacia Biothech Inc., Uppsala, Sweden), 1.25 mM dNTP (Boehringer-Mannheim, Mannheim, Germany) and 200 U M-MLV reverse transcriptase (Gibco BRL). PCR was performed using 0.25 mM dNTP, 0.25 U Taq polymerase (Perkin Elmer, Norwalk, CT, USA), 10 pmole primer pair, and 3 µL cDNA with a thermal cycler (Perkin Elmer). The primer sequences were as follows: PPARγ, 5'-TCCGTGATGGAAGACCACTC-3' (sense, 190–209) and 5'-CCCTTGCATCCTTCACAAGC-3' (antisense, 502–521; GeneBank U09138); and β-actin, 5'-TTGTAACCAACTGGGAC GATTGG-3' (sense, 1552–1575) and 5'-GATCTTGATCTTCATG GTGCTAGG-3' (antisense, 2967–2991; GeneBank J00691). PCR cycling conditions were as follows: 92℃ for 30 sec, 55℃ for 30 sec, and 72℃ for 1 min.
In vitro PDT using PPME and apoptosis assay
For photodynamic analysis, target cells (1×106 in a 35-mm2 dish) were pre-incubated with 1.5 mL of RPMI added with varied doses of PPME (0.1–1 µg/mL) for 1 h at 37℃. Following incubation, the cells were exposed to light. The light source was a 200-W halogen lamp (MVI, Micro Video Instruments Inc., Avon, MA, USA) attenuated by a 515-nm filter. The total power output for the irradiation of the cells was adjusted to 120 mJ/cm2 as indicated by a Laser power meter (Metrologic Instruments, Inc., Blackwood, NJ, USA). Then, cells treated with PPME/PDT were further incubated at 37℃ for 24 h. Cells were stained with propidium iodide (PI) and annexin V-FITC (Biosource International, Camarillo, CA, USA), and cell death was analyzed via flow cytometry using the FACScan flow cytometer (Becton-Dickinson, CA, USA).
MTT assay
Cells (1×103 to 3×103) were cultured in 96-well plates with or without troglitazone for various concentrations. After incubation (1, 2, 4, and 6 days respectively), cells were washed in phosphate-buffered saline (PBS, pH 7.4), added to 50 µL of the 1-mg/mL MTT solution in sterile PBS, and incubated for 4 h at 37℃. Then, the plate was centrifugated at 3000 rpm for 10 min, supernatant was carefully removed, and remaining adhered cells were dissolved in 50 µL DMSO (Sigma). The plate was read at 570 nm using an ELISA reader (Softmax Pro, Biorad, Hercules, CA, USA).
Western blot analysis
Cells were cultured in 6-well plates with or without 50 µM troglitazone and PDT and then lysed with lysis buffer containing 50 mM Tris-HCl (pH 7.4), 100 mM NaCl, 0.5 mM EDTA, 0.2% NP-40, and 2 mM MgCl2 supplemented with protease inhibitor cocktail (Sigma). The samples were analyzed via 10% SDS-polyacrylamide gel electrophoresis. After transfer, membranes were blocked with 5% non-fat dry milk in PBS (w/vol)-0.05% Tween 20 and incubated overnight at 4℃ with anti-caspase-3 Ab (Transduction Lab., Lexington, KY, USA), anti-cytochrome-C Ab (Pharmingen, San Diego, CA, USA), and anti-α-tubulin Ab (Oncogene, Boston, MA, USA). Horseradish peroxidase-conjugated goat anti-mouse antibody (DAKO, Glostrup, Denmark) was used at a 1:1000 dilution as the secondary antibody, and reactive proteins were detected by incubation in SuperSignal substrate (Pierce, Rockford, IL, USA) followed by exposure to X-ray film (Eastman Kodak Company, Rochester, NY, USA). For quantification, densities of procaspase-3 bands were normalized to the density of cytochrome C bands measured using Image J software (developed by Wayne Rasband, National Institutes of Health, Bethesda, MD, USA; available at http://rsb.info.nih.gov/ij/index.html).
RESULTS
Troglitazone inhibits the proliferation of DLD-1 cells
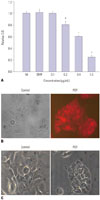
To test the mRNA expression of PPARγ in DLD-1 cells, we performed RT-PCR. Consistent with previous reports,21 PPARγ mRNA was detected in DLD-1 (Fig. 1). To test whether troglitazone influences the proliferation of DLD-1 cells, an MTT assay was performed. As shown in Fig. 1, the PPARγ agonist, troglitazone, had almost no effect on the growth of DLD-1 cells during the first 2 days of the culture, although it significantly inhibited the expansion of DLD-1 cells after 4 days in a troglitazone dose-dependent manner. A dose of 100 µM troglitazone reduced cell survival and induced a morphological change of DLD-1 (data not shown). The number of cells recovered from the group treated with 50 µM troglitazone on day 6 was threefold less than that of the control, suggesting that troglitazone effectively blocks the proliferation of DLD-1 cells.
PDT reduces the viability of DLD-1
To test the effects of PDT on the cell viability of DLD-1, cell viability was measured via MTT assay 24 h after PDT with various concentrations of PPME. PDT reduced the viability of DLD-1 in a PPME dose-dependent manner (Fig. 2A). To determine the photosensitizer uptake in the cells, DLD-1 cells were incubated with 200 nM PPME for 1 h, and the uptake of PPME was detected via fluorescent microscopy. Fig. 2B showed a strong uptake of photosensitizer in the cytoplasm of DLD-1 cells. Using a light microscope, we observed floating and membrane blebbing of DLD-1 after 0.4 µg PPME/PDT, indicating that PPME/PDT induced the cellular change of apoptosis (Fig. 2C).
Troglitazone enhances PDT-induced cell death
To test whether PPARγ enhances PDT-induced apoptosis, we examined the effect of PPARγ activation on the death of tumor cells using troglitazone and PDT. DLD-1 cells were pre-incubated with 10 mM troglitazone for 24 h and then treated with PDT. As shown in Fig. 3A, 0.4 µg/120 mJ PDT reduced cell viability by approximately 60%. This effect was augmented by pre-incubating cells with 20 mM troglitazone for 24 h. On using HCT-15 and HT-29 cells, we also observed a similar result of decreased cell viability in response to combination treatment (Fig. 3B). These results suggest that the PPARγ ligand, troglitazone, enhances the sensitivity of colon cancer cells to PDT-induced cell death.
Combination treatment induces apoptosis
PDT can induce either apoptotic or necrotic cell death. Generally, high-dose PDT induces cancer cell necrosis immediately, while low doses show delayed-type apoptosis, rather than necrosis. In our system, cells treated with over 0.5 µg/mL photosensitizer underwent necrotic cell death. To test whether combination treatment induces apoptosis or necrosis, we performed a flow cytometric analysis using the Annexin V/PI staining method, which is commonly used to differentiate necrotic and apoptotic cell populations. As shown in Fig. 4, troglitazone plus PDT showed a drastic increase in apoptotic cell population compared to other treatments.
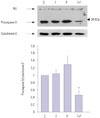
Combination treatment induces procaspase-3 degradation
Caspase activation is an important event in the apoptotic pathway and can serve as a marker of apoptosis. Given that caspase-3 is a central executioner caspase, we performed a Western blot analysis with anti-caspase-3 antibody. As shown in Fig. 5, procaspase-3 was reduced in the combination treatment with PDT plus troglitazone. This result suggests that the combination of PDT and troglitazone significantly induces the degradation of procaspase-3 and leads to apoptosis, in agreement with data in Fig. 4. It is tempting to speculate, therefore, that the combination of PDT and troglitazone is a potent killing modality in DLD-1.
DISCUSSION
PDT induces apoptosis in a variety of tumor cell lines yet does not induce apoptosis among most normal cells. It has been clinically applied in treatments of head and neck, brain, bladder, skin, and intrathoracic malignancies.1 PPARγ ligands inhibit the proliferation of colon cancer cells and induce apoptotic or non-apoptotic cell death in various tumors. In this study, we investigated the effect of the PPARγ ligand troglitazone on the efficiency of PDT in DLD-1 colon cancer cells. Our data showed that troglitazone not only inhibited the proliferation but also sensitized DLD-1 to PDT-induced cell death. Whereas no direct cytotoxicity was observed in low-dose treatments of troglitazone (<50 µM), high-dose treatments resulted in a toxic effect on DLD-1 cells. The effect of PDT is presented in Fig. 3 and we selected a PDT dose of 0.4 µg/120 mJ for IC50. Using a combination of PDT and low-dose troglitazone, we minimized immediate necrotic cell death. Although no inhibitory effect was shown in a single treatment, the combination of PDT with troglitazone effectively killed DLD-1 cells. DLD-1 underwent apoptosis rather than necrosis, as shown in Fig. 4, and caspase-3 might also be involved in PDT/troglitazone-induced apoptosis (Fig. 5).
Although PDT has tumoricidal activity, the cellular mechanism responsible for the tumor selectivity was not completely clarified. As a tumor killing mechanism, it is reported that PDT invokes the production of reactive oxygen species (ROS), and this kills tumor cells successfully. 222324 Other data indicate that certain kinds of photosensitizers directly target mitochondria, inducing apoptosis by releasing cytochrome C and activating various caspases.252627 Recently, Matroule, et al.28 reported that HCT-116 underwent apoptosis via PDT and that mitochondria and ROS played a central role in PPME-mediated apoptosis. Tumoricidal activity of PDT has also been reported in many types of cancer cell lines as well as colon cancer cell lines.26 According to a previous report, PDT induces activation of NF-κB as well as an apoptotic pathway.28 It was suggested that the activation of NF-κB may involve restoration from a death pathway after PDT. Among the various effects of PPARγ ligands, they are particularly known to antagonize the activities of several transcription factors including AP-1, STAT, and NF-κB.293031 Our data demonstrate that combination of PDT and troglitazone increases the degree of apoptosis and degradation of procaspase-3, raising the possibility that troglitazone effectively inhibits the activation of the survival pathway after PDT.
In addition, it has been reported that PPARγ regulates the expression of cyclin-dependent kinase inhibitors p18 and p21, inducing growth arrest and differentiation.3233 It may be possible that pretreatment of troglitazone may increase the susceptibility to PDT via disturbance in cell cycle regulation. However, the sensitizing mechanism of troglitazone in PDT-induced apoptosis remains to be determined.
PDT induces apoptosis as well as necrosis in cancer cells. Our data demonstrate for the first time that troglitazone treatment enhances PDT-induced cytotoxicity in DLD-1 cells, and these results suggest a new therapeutic tool for combination treatment that is free of severe side effects. However, it must be further investigated whether a combination of PDT with PPARγ agonist is safe in animals, and many experiments will be required. In summary, our study resulted in a novel observation of troglitazone sensitizing PDT-induced apoptosis, which indicates that this combination may be an effective modality for the treatment of colon cancer.



 XML Download
XML Download