

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The endothelium plays a pivotal role in the maintenance of vascular homeostasis by modulating vascular tone, leukocyte adhesion, platelet aggregation, smooth muscle cell proliferation, and migration. Recent discoveries have revealed that endothelial nitric oxide (NO) is an important vasoprotective molecule contributing to endothelial functions, and impaired NO signaling associated with endothelial dysfunction is now regarded as an early marker of cardiovascular diseases.
Endothelial arginase inhibits endothelial NO synthase (eNOS) activity by depleting its common substrate (L-arginine), which lowers NO bioavailability and leads to vascular diseases in aging, hypertension, and atherosclerosis.1234 In atherogenic-prone apolipoprotein E-null (ApoE-/-) mice, arginase inhibition restores NO and reactive oxygen species (ROS) balance, endothelial function, and arterial stiffness to the wild-type phenotype.5 In the aging cardiovascular system, inhibition of S-nitrosylation-dependent arginase I activation restores depressed endothelial function.6 Furthermore, we demonstrated that in both pathophysiological and normal physiological vasculature, arginase inhibition induced eNOS activation through phosphorylation at Ser1177.7
Redox-imbalance may result in the generation of peroxynitrite, which is associated with cardiovascular diseases as it initiates the modification of DNA, carbohydrates, and proteins. Several protein molecules including eNOS and arginase are affected by post-translational modifications such as peroxynitrite-induced S-nitrosylation, which is involved in the regulation of enzyme activity. In endothelial cells, peroxynitrite induces the S-nitrosylation of eNOS and the oxidation of the essential cofactor for NOS, (6R-)5,6,7,8-tetrabiopterin (BH4), resulting in impaired endothelial function through NOS uncoupling.8 In addition, peroxynitrite also reduces intracellular L-arginine concentration by decreasing endothelial L-arginine influx and increasing L-arginine efflux via nitrosylation of cationic amino acid transporter. The decline in L-arginine concentration results in less NO production.9
Although a peroxynitrite generator, 5-amino-3-morpholinyl-1,2,3-oxadiazolium chloride (SIN-1), induces the upregulation of arginase activity and expression contributing to endothelial dysfunction,10 it is not completely known whether arginase inhibition and L-arginine supplement could restore peroxynitrite-dependent endothelial dysfunction and if so, which mechanism is involved. Therefore, with the hypothesis that peroxynitrite-dependent endothelial dysfunction may be induced by L-arginine depletion, which was derived from arginase activation and further provoked eNOS uncoupling, we tested whether increased L-arginine bioavailability to eNOS via arginase inhibition and L-arginine supplement could restore eNOS activation and depressed endothelial function. In our study, we demonstrated that stimulation of human umbilical vein endothelial cells (HUVECs) with peroxynitrite increased arginase activity with an effect on expression levels of arginase I/II. Increased arginase activity was associated with eNOS uncoupling. Arginase inhibition and L-arginine supplement enhanced the phosphorylation of Ser1177, reduced the phosphorylation of Thr495, induced the augmented stability of eNOS dimers, and reciprocally regulated NO production and ROS generation. Furthermore, impaired vascular reactivity caused by peroxynitrite preincubation was restored by treatment with an arginase inhibitor. These findings suggest a new vasoprotective effect of arginase inhibition during nitroso-redox imbalance.
MATERIALS AND METHODS
Materials
ABH [2(S)-amino-6-boronohexanoic acid], SIN-1 (Amino-3-morpholinyl-1,2,3-oxadiazolium chloride), ONOO-, and FeTPPS [5,10,15,20-Tetrakis (4-sulfonatophenyl) prophyrinato iron (III)] were purchased from Calbiochem (Darmstadt, Germany). All other chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated.
Cell culture
HUVECs were purchased from Cascade Biologics (Portland, OR, USA) and maintained in Medium 200 containing low serum growth supplement according to the supplier's protocol.
Arginase activity measurement
Arginase activity was measured by determination of urea content using α-isonitrosopropiophenone as described previously.3 Briefly, supernatants of extracted cell lysates were prepared by incubation with lysis buffer (50 mM Tris-HCl, pH7.5, 0.1 mM EDTA, and protease inhibitors) for 30 min at 4℃ followed by centrifugation at 14000× g for 20 min at 4℃. HUVECs were treated with arginase inhibitor (ABH) before SIN-1 stimulation. Assays on aortic vessel samples were performed following homogenization in lysis buffer.
NO measurement
A NO assay kit (Calbiochem) was used to measure nitrite/nitrate (NOx) via the Griess reaction, after conversion of nitrate to nitrite with nitrate reductase. The concentration of NOx from cell lysates was expressed as µmol/g protein.
Measurement of NO release and ROS production with fluorescent dyes DAF and DHE
All experimental procedures using mice were approved by the Institutional Review Board at Kangwon National University. Aortic rings from 10-week-old male wild-type C57BL/6J mice were prepared for arginase activity assays or labeled with dihydroethidine (DHE, 1 µmol/L, 5 min) or 4-amino-5-methylamino-2',7'-difluorescein (DAF, 5 µmol/L, 5 min), fluorescent probes for superoxide and NO, respectively. Images were acquired using an Olympus BX51 epifluorescence microscope. Fluorescent intensity was measured as previously described5 using MetaMorph software (Molecular Devices, Inc., Sunnyvale, CA, USA).
Western blot analysis and eNOS dimer blot analysis
Cells were lysed in SDS-sample buffer (62.5 mMTris, pH 6.8, 2% SDS, and 10% glycerol) and then sonicated for 5 s to reduce sample viscosity. Each sample was resolved by 10% SDS-polyacrylamide gel electrophoresis (SDS-PAGE), transferred to polyvinylidene difluoride (PVDF) membrane (BioRad, Hercules, CA, USA), analyzed with antibodies [eNOS, inducible nitric oxide synthase (iNOS), hArg1, hArg2 antibodies from Santa Cruz Biotechnology and phospho-eNOS Ser1177 and phosphoeNOS Thr495 antibodies from BD Biosciences] according to the suppliers' protocols, and visualized with peroxidase and an enhanced-chemiluminescence system (Thermo Scientific Pierce, Waltham, MA, USA). Normalization was performed using an anti-β-tubulin antibody (BD bioscience, 1:1000). A densitometry analysis of bands was performed using NIH ImageJ. Dimers and monomers of eNOS were separated by low-temperature SDS-PAGE and analyzed as described above.
Vascular tension assay
Male wild-type (WT, C57BL/6J) mice were anesthetized using isoflurane, and the thoracic aortic vessel was rapidly removed. Each aorta was placed on ice-cold oxygenated Krebs-Ringer bicarbonate solution (NaCl 118.3, KCl 4.7, MgSO4 1.2, KH2PO4 1.2, CaCl2 1.6, NaHCO3 25, glucose 11.1 mM) and cleared of connective tissue. Each mouse aorta was cut into 1.5-mm rings and suspended between two wire stirrups (150 µm) in a myograph (Multi myograph system DMT-620) containing 10 mL Krebs-Ringer solution (95% O2-5% CO2, pH 7.4, 37℃). One stirrup was connected to a three-dimensional micromanipulator, and the other stirrup was connected to a force transducer. The rings were passively stretched at 10-minute intervals in increments of 100 mg in order to reach optimal tone (600 mg). After the arterial rings were stretched to their optimal resting tone, the contractile response to 100 mMKCl was determined. The response to a maximal dose of KCl was used to normalize the responses to agonist across vessel rings. A dose response (10-9–10-5 M) to the vasoconstrictor, U46619, was analyzed, followed by a dose response (10-9–10-5 M) to the vasodilator, acetylcholine (Ach), after preconstriction with U46619 (10-8 M). To confirm that the vasorelaxation activity was NO-dependent, the endothelial layer was physically removed by gentle rubbing.
Determination of intracellular L-arginine concentrations
The intracellular concentration of L-arginine was determined via high-performance liquid chromatography (HPLC) using pre-column derivatization with o-phthalaldehyde (OPA) according to a modification of a previously published method.11 L-arginine (100 µmol/L) was added to the cell lysate (0.5 mL) as an internal standard. The samples were extracted on solid-phase extraction cartridges (CBA Bond elute, Varian). Recovery rates were 87.5±3.9%. Eluates were dried over nitrogen and resuspended in double-distilled water for HPLC analysis. HPLC was performed on a computer-controlled Waters chromatography system (M600E) consisting of an automatic injector (M7725i, Waters Co., Victoria, Australia) and a fluorescence detector (FP-1520, Jasco Co., Queensland, Australia) in the Central Laboratory of Kangwon National University. Samples were incubated for exactly 1 minute with OPA reagent (5.4 mg/mL OPA in borate buffer, pH 8.4, containing 0.4% 2-mercaptoethanol) before automatic injection into the HPLC. The OPA derivative of L-arginine was separated on a 150×4.6 mm (3.5 µm) Zorbax Eclipse XDB-C18 column with a fluorescence detector set at Ex 340 nm and Em 450 nm. Samples were eluted from the column with 0.96% citric acid/methanol (70:30, pH 6.8) at a flow rate of 1.5 mL/min.
RESULTS
SIN-1-derivied peroxynitrite increased arginase activity/expression and reduced NO generation in endothelial cells without changing eNOS protein level
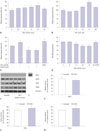
First, we measured arginase enzyme activity in SIN-1-treated HUVECs. SIN-1 treatment induced a time-dependent increase in arginase activity (Fig. 1A, *p<0.05 vs. untreated), with a maximal effect at 18 hours of 50 µM SIN-1 (127.4±5.0%). In addition, the different concentration of SIN-1 at 18 hours increased arginase activity in a dose-dependent manner (Fig. 1B, *p<0.01 vs. untreated) with a maximal effect at 50 µmol/L of SIN-1 (135.7±3.8%). As reactivities of major products of SIN-1 (NO, superoxide, and peroxynitrite) are relatively high, it is possible that the effect of SIN-1 is highly dependent on time and concentration. Hence, the optimal concentration (50 µmol/L) and treatment time (18 hours) for SIN-1 were used in all subsequent experiments unless otherwise stated.
As SIN-1 releases equimolar NO and superoxide, which react to form peroxynitrite,12 we next tested whether the effect of SIN-1 is dependent on peroxynitrite generation. HUVECs were preincubated with 5 µM FeTPPS, a decomposition catalyst of peroxynitrite, followed by stimulation with SIN-1. FeTPPS blocked the increased arginase activity induced by SIN-1 stimulation (Fig. 1C, *p<0.01 vs. untreated; #p<0.01 vs. SIN-1 alone). Peroxynitrite (50 µmol/L, 18 hours) also significantly increased arginase activity (Fig. 1C, *p<0.01 vs. untreated). These data indicate that SIN-1-derived peroxynitrite increased arginase activity.
The increased arginase activity was associated with a decrease in NO metabolites (nitrite and nitrate). As demonstrated in Fig. 1D, SIN-1 stimulation resulted in a time-dependent decrease in NOx production after 12 hours of SIN-1 treatment (Fig. 1D, *p<0.01 vs. untreated). Moreover, peroxynitrite decomposition with FeTPPS (5 µM) also prevented a SIN-1-dependent decrease in NOx production (Fig. 1D, #p<0.01 vs. SIN-1 alone for 18 hours, 97.8±1.3% vs. 80.3±1.1%).
As SIN-1 induced an increase in arginase enzyme activity and a decrease in NOx production, we wished to determine if protein expression levels were changed. As shown in Fig. 1E and F, protein abundance of eNOS was not affected, and iNOS was not detected. However, the expression of arginase I and II proteins increased in SIN-1-treated HUVECs for 18 hours (Fig. 1G and H, *p<0.05 vs. untreated).
eNOS phosphorylation and dimer stability by SIN-1 were restored by preincubation with an arginase inhibitor and L-arginine supplement
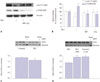
Given that SIN-1 stimulation decreased NOx production without affecting eNOS abundance, we wished to determine the molecular mechanism underlying this phenomenon. It is reported that eNOS increases enzymatic activity when the Ser1177 site is phosphorylated and the Thr495 site is de-phosphorylated. As shown in Fig. 2A and B, SIN-1 treatment significantly reduced phospho-eNOS Ser1177 and increased phospho-eNOS Thr495 (Fig. 2B, *p<0.05 vs. untreated). However, arginase inhibition and L-arginine supplement markedly reversed the effect of SIN-1 on eNOS phosphorylation (Fig. 2B, #p<0.05 vs. SIN-1 alone, ##p<0.01 vs. SIN-1 alone).
We next determined the effects of SIN-1 on the stability of eNOS dimers. As shown in Fig. 2C, SIN-1 induced a significant decrease in the abundance of eNOS dimers (Fig. 2C, *p<0.05 vs. untreated, 1.28±0.06 vs. 1.49±0.05). On the other hand, preincubation with ABH and L-arginine before SIN-1 treatment significantly enhanced the stability of eNOS dimers (Fig. 2D, *p<0.01 vs. SIN-1 alone, 1.89±0.08 vs. 1.38±0.05; #p<0.01 vs. SIN-1 alone, 1.93±0.06 vs. 1.38±0.05). These data suggest that an increased intracellular L-arginine concentration may play a major role in eNOS coupling in peroxynitrite-dependent pathological conditions.
SIN-1-induced decrease in NO generation and increase in ROS production that were restored by incubation with arginase inhibitor and L-arginine in isolated mouse aortas
With the hypothesis that arginase inhibition and intracellular L-arginine amount are involved in SIN-1-dependent regulation of eNOS activity, we next determined changes in NO and ROS production in the endothelium of isolated mouse aortas using the fluorescent dyes DAF and DHE (see Materials and Methods).5 Incubation of aortic rings with SIN-1 decreased the DAF fluorescence slope (Fig. 3A, *p<0.01 vs. untreated, 0.69±0.08 vs. 1.28±0.34 fluorescence intensity/second). Remarkably, arginase inhibition (ABH, 10 µM) restored the DAF fluorescence rate (Fig. 3A, #p<0.01 vs. SIN-1 alone, 1.41±0.40 vs. 0.69±0.08 fluorescence intensity/second). To confirm the effects of L-arginine supplement on eNOS phosphorylation and dimer stability (Fig. 2), we directly added L-arginine (10 mM) to SIN-1-treated aortic rings and then performed a similar DAF bioassay. The DAF fluorescence rate decrease induced by SIN-1 was markedly attenuated in L-arginine-treated aortic endothelium (Fig. 3A, #p<0.01 vs. SIN-1 alone, 1.24±0.15 vs. 0.69±0.08 fluorescence intensity/second). However, the increased slope of DAF fluorescence by ABH incubation did not change with treatment of iNOS inhibitor (1400W, data not shown). Taken together, these data suggest that the SIN-1-induced decrease in endothelial NO production may be dependent on arginase activity affecting the intracellular L-arginine level. NOS inhibitor, NG-Nitro-L-arginine methyl ester (L-NAME, 100 µM) resulted in almost complete reduction of the slope of DAF fluorescence to 0.
To determine whether arginase activation by SIN-1 was associated with increased ROS production via eNOS uncoupling, we measured O2•- production using DHE in isolated aortas treated with L-NAME (100 µM), ABH (10 µM), L-arginine (10 mM), or all three. There was a significant increase in ROS production in the endothelium of SIN-1 treated aortas (Fig. 3B, *p<0.01 vs. untreated, 2.54±0.97 vs. 1.02±0.33 fluorescence intensity/second). Consistent with this data, ROS production was significantly decreased with ABH treatment (Fig. 3B, #p<0.01 vs. SIN-1 alone, 1.02±0.29 vs. 2.54±0.97 fluorescence intensity/second) and L-arginine incubation (Fig. 3B, #p<0.01 vs. SIN-1 alone, 1.02±0.32 vs. 2.54±0.97 fluorescence intensity/second). Interestingly, incubation of SIN-1-treated aortic rings with L-NAME prevented a SIN-1-induced increase in ROS production, and pretreatment with ABH or L-arginine resulted in a slope similar to the DHE response in untreated aortas (Fig. 3B, NS: non-significant among the groups). These results suggest that enhanced ROS production under peroxynitrite-dependent stress condition is due to eNOS uncoupling.
Arginase inhibition restored intracellular L-arginine concentration
We also measured the intracellular L-arginine concentration in HUVECs. Under control conditions, the intracellular content of L-arginine was 100±5.0% (170±8.5 µM L-arginine), and the L-arginine content fell to 82.8±4.8% after exposure to SIN-1 (Fig. 4, *p<0.01 vs. untreated). However, preincubation with ABH maintained L-arginine content at 96.9±2.8% (Fig. 4, #p<0.01 vs. SIN-1 alone).
Arginase inhibition restored impaired vessel reactivity induced by SIN-1 in an endothelium-dependent manner
Considering that both arginase inhibition and L-arginine restored the SIN-1-induced decrease in vascular NO production, we determined whether vascular function was altered in SIN-1 treated aortas. First, we determined the vascular response to KCl. SIN-1 treatment did not significantly change the maximal tension development in response to 60 mMKCl (Fig. 5A, NS: non-significant between untreated vs. SIN-1, 1.60±0.12 g vs. 1.45±0.15 g).
The vasoconstriction response to the agonist, U-46619, was markedly enhanced in SIN-1-treated aortas, as manifest by a leftward shift in the dose-response curves, an increase in Emax (Fig. 5B, *p<0.01 vs. untreated, 241.6±5.0% vs. 209.3±5.2%), and a reduction in log EC50 (Fig. 5B, *p<0.01 vs. untreated, -7.92±0.05 M vs. -7.76±0.05 M). However, incubation with ABH for 18 hours significantly reduced Emax (Fig. 5B, #p<0.01 vs. SIN-1 alone, 172.0±6.6% vs. 241.6±5.0%) and increased log EC50 (Fig. 5B, #p<0.01 vs. SIN-1 alone, -7.43±0.07 M vs. -7.92±0.05 M). Moreover, peroxynitrite decomposition with FeTPPS (5 µM) also reduced the Emax of the SIN-1-treated aortas to the same level as the Emax of untreated aortas (Fig. 5B, SIN-1+FeTPPS vs. untreated, 201.7±5.3% vs. 209.3± 5.2%, non-significant).
To determine the effects of arginase inhibition on endothelial-dependent vasorelaxation, mouse aortas were preconstricted with U46619 (10-8 mol/L), and dose-response curves were constructed in response to the endothelial-dependent vasodilator, Ach, and the endothelial-independent NO donor, sodium nitroprusside (SNP). Ach resulted in significant dose-dependent relaxation in the mouse aortas. The vasorelaxation responses to SIN-1 in the aortic rings were significantly attenuated compared with those from untreated controls. The Emax was 85.9±2.3% vs. 90.2±2.5% (Fig. 5C, *p<0.05 vs. untreated), and the log EC50 was -7.10±0.07 M vs. -7.48±0.10 M (Fig. 5C, *p<0.05 vs. untreated). FeTPPS (5 µM) increased the Emax of SIN-1-treated aortas to the untreated control level (Fig. 5C, SIN-1+FeTPPS vs. untreated, 93.2±1.8% vs. 90.2±2.5%, non-significant). Surprisingly, the Emax in vessel rings treated with SIN-1 in the presence of ABH were enhanced compared to those of SIN-1 treated rings (Fig. 5C, #p<0.01 vs. SIN-1 alone, 109.5±2.6% vs. 85.9±2.3%); the log EC50, however, was not significantly changed (Fig. 5C, SIN-1 vs. SIN-1+ABH, -7.10±0.07 M vs. -7.09±0.06 M). De-endothelialized rings did not response to Ach (Fig. 5C). In contrast, vessels preconstricted with U46619 did not show a significant change in cumulative dose responses to SNP among the groups (Fig. 5D).
DISCUSSION
First, the concentration of peroxynitrite (Calbiochem, Feltham, UK) was determined spectrophotometrically based on an extinction coefficient of 1670 M-1 cm-1 at 302 nm in 1.5 M NaOH. We quantified peroxynitrite generated from SIN-1 decomposition by quenching the NADH fluorescence, which has been reported to correspond to the conversion of NADH to NAD+ by peroxynitrite.13 NADH fluorescence was measured with an excitation wavelength of 340 nm and an emission wavelength of 460 nm (LS-5B, Perkin-Elmer Instruments, Waltham, MA, USA). SIN-1 treatment at 50 µM, which was used in all experiments, generated peroxynitrite at a concentration of 0.65 nM over 30 min.
NO is a vasoprotective molecule that has led to the development of therapies to enhance NO signaling, including dietary L-arginine,1415161718 drug-eluting stents,19 inhalational NO gas,2021 NOS gene therapy,2223 and recently arginase inhibition.1524 Unfortunately, gene therapy remains limited due to problems with the delivery system and vectors, as well as mutational concerns. Inhalational NO gas has a limited use in terms of dose, exposure, and systemic toxicity.2526 Pharmacological NO donors may contribute to vascular injury prior to initiation of therapy due to surgical manipulation. Moreover, oral L-arginine therapy generated controversial results.151617 On the other hand, inhibition of arginase, a novel therapeutic target, actively augments NO production, and has beneficial effects on cardiac and vascular wall function in atherogenesis, diabetes, and aging.1452427 Our data support the hypothesis that arginase inhibiton, as well as L-arginine supplementation, has protective effects on endothelial dysfunction and hypertension.
The activation and expression of endothelial arginase can be induced by a variety of vascular insults, although the dominance of arginase I and II expression may depend on the organ, disease state, and species.28 Angiotensin II is known to elevate arginase activity through the induction of arginase I via RhoA/RhoA activated kinase (ROCK) activation.29 Interestingly, arginase I activity was increased by stabilization of the arginase I trimer via S-nitrosylation at Cys303,6 which required direct interaction with the oxygenase domain of iNOS.30 In the case of arginase II, the enzymatic activity was increased in endothelial cells by incubation with thrombin31 and oxidized low-density lipoprotein (oxLDL).3 These agonists, signaling through RhoA-dependent ROCK activation, activate arginase II without effecting arginase protein expression. Taken together, it seems that activation of the RhoA/ROCK pathway can elevate arginase expression and activity. Interestingly, a previous study reported that the RhoA molecule also contains a redox-active cysteine (Cys18), which ROS and reactive nitrogen species (RNS) may target. ROS and RNS, particularly the superoxide anion radical (O2•-) and nitrogen dioxide (•NO2), can enhance GDP release from RhoA and thus modulate RhoA activity.32
Peroxynitrite is a highly reactive oxidant formed by the reaction of superoxide and NO33 and is an indicator of the redox state. Increased formation of peroxynitrite, referred to as redox imbalance, has a wide array of tissue-damaging effects in vivo, including lipid peroxidation, inactivation of enzymes and ion channels via protein oxidation and nitration, and inhibition of mitochondrial respiration.33 Peroxynitrite has been detected in disease conditions such as diabetes, ischemia-reperfusion, sepsis, inflammation, and atherosclerosis.343536 Endothelial NOS may play an important role in the regulation of redox balance in endothelial cells, as ROS and NO production are dependent on the stability of the eNOS dimer. Several mechanisms could explain the uncoupling of eNOS to produce predominantly ROS in pathophysiological conditions including: 1) substrate (L-arginine) depletion, 2) cofactor (BH4) depletion, 3) loss of dimerization, and 4) altered eNOS phosphorylation. Among them, peroxynitrite can induce eNOS uncoupling via BH4 depletion (an oxidation reaction in BH2), disruption of the eNOS-heme complex,37 and oxidation of the zinc-thiolate complex.38 Peroxynitrite can also nitrosylate the cationic amino acid transporter, resulting in decreased endothelial L-arginine transport and an increased rate of arginine efflux, thus reducing intracellular L-arginine concentration and negatively effecting NO production.39 Furthermore, increased iNOS-dependent NO can induce increased arginase I enzyme activity by S-nitrosylation leading to decreased intracellular L-arginine bioavailability.6 These effects of peroxynitrite might explain why chronic treatment for 18 hours with SIN-1 resulted in activation of arginase (Fig. 1A, B, and C) and decreased NO production (Fig. 1D and 3A), although eNOS expression did not change (Fig. 1E).
Reduced intracellular BH4 level has been demonstrated as a cause of endothelial NOS uncoupling in experimental vascular disease states. It is also reported that decreased L-arginine bioavailability alone is sufficient to uncouple NOS.40 Interestingly, preincubation with arginase inhibitor or L-arginine prevented peroxynitrite-dependent eNOS uncoupling (Fig. 2D). This result is consistent with a previous report showing that inhibition of arginase prevented the decrease in eNOS dimers in endothelial cells after treatment with oxLDL, and the effect of arginase on NOS uncoupling was independent of vascular BH4 concentration.5 Furthermore, our data suggest that increased arginase activity is associated with eNOS uncoupling, and enhancement of the L-arginine amount may play a critical role in maintaining the stability of eNOS dimers through increased eNOS phosphorylation at Ser1177 and decreased phosphorylation at Thr495, which contribute to the restoration of the nitroso-redox balance in endothelial cells (Fig. 5). Hence, arginine inhibition and L-arginine supplementation are able to reverse the effects of peroxinitrite on endothelial cells, as shown in our data.
In conclusion, peroxynitrite stimulation induced an increase in arginase activity and reciprocally regulated NO production that was restored by increasing the L-arginine level. The increased L-arginine level was associated with eNOS dimerization via eNOS phosphorylation at Ser1177. Furthermore, arginase inhibition and L-arginine supplementation attenuated peroxynitrite-induced vascular dysfunction by a mechanism involving increased stability of eNOS dimers.



 XML Download
XML Download