

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The main functions of the testes are testosterone production and spermatogenesis. These two functions are controlled by the hypothalamus-pituitary-gonad axis. Gonadotropin-releasing hormone (GnRH) from the hypothalamus stimulates the production of follicle-stimulating hormone (FSH) and luteinizing hormone (LH) in the anterior pituitary.1 FSH binds to its receptor on the surface of Sertoli cells to regulate spermatogenesis.23 LH binds to its receptor on the Leydig cell membrane to stimulate testosterone production. The testosterone produced by LH negatively regulates GnRH production in the hypothalamus.456
There are four types of forkhead box class O (Foxo) transcription factors: Foxo1 (FKHR, forkhead in rhabdomyosarcoma), Foxo3 (FKHRL2, FKHR-like1), Foxo4 (AFX, acute-lymphocytic-leukemia-1), and Foxo6. These Foxo proteins regulate stress responses, aging, insulin sensitivity, and ontogenesis,78 and their transcription is inhibited by phosphoinositide 3-kinase (PI3K). PI3K signaling phosphorylates AKT, which then phosphorylates Foxo3 at Ser24, Thr32, and Ser56 residues. These phosphorylated sites recruit 14-3-3 protein to guide Foxo3 from the nucleus into the cytoplasm. Finally, Foxo3 is removed by proteasomes.9
In female reproduction, Foxo1, Foxo3, and Foxo4 are expressed in the granulosa cells at various stages of follicle development.10 Foxo1 in granulosa cells inhibits cyclin D2 gene expression and increases the nuclear localization of p27kip proteins, which makes it a key regulator of G1/S transition. Foxo proteins also play an important role in regulating ovarian function by pituitary gonadotropins.1011 In a previous study, Foxo3-null female mice exhibited age-dependent fertility issues and were completely sterile at 10 weeks or older. In Foxo3-/- ovaries at 9.5 weeks, oocytes in developing follicles appeared to have degenerated, reflecting atretic change. At 12 weeks, Foxo3-/- ovaries had no developing follicles. These indicated that Foxo3 is important in ovarian follicular development.111213 Meanwhile, in males, germ line specific Foxo1 KO mice showed defective proliferative expansion and small testes, which was not due to cell death, but rather to renewal of spermatogonial stem cells.14 However, the function of Foxo3 in Leydig cells is not clear.15
Foxo3 is important not only in females, but also in males. Foxo3 expression and location are likely to be dynamic throughout life. In this study, Foxo3 expression and location were investigated from mouse embryonic stage to 12 weeks, and the role of Foxo3 in Leydig cells was investigated to outline the function and regulation of Leydig cells.
MATERIALS AND METHODS
Animals and testis preparation
C57/BL6 (Jackson Labs, CA, USA) male mice were housed in a barrier facility under normal light and dark conditions and fed ad libitum. Testes were isolated at postpartum days (PPD) 1 and 5 and postpartum weeks (PPW) 3, 4, 5, and 12. Testes were removed, fixed in 10% formalin, and embedded in paraffin. All procedures were approved by the Animal Care and Use Committees at Yonsei University College of Medicine and Northwestern University.
Plasmid and adenovirus construction
The mStARp-Luc plasmid was constructed by inserting the mouse steroidogenic acute regulatory (StAR) promoter (2730 bps) into the multiple cloning site of pGL3 basic vector. As synthetic poly A (spa) region in PGL3 basic vector contained two FOXO binding sequences, we removed them using NotI and KpnI.16 The human FOXO3 triple mutant (FOXO3-TM) was generated by substituting Thr32, Ser253, and Ser315 with alanine residues. This FOXO3-TM cannot be phosphorylated by Akt and is constitutively activated.1718 Recombinant adenoviral vectors carrying FOXO3 [wild type (WT) or TM] were generated as previously described.8 Adenovirus carrying β-galactosidase (Ad-Gal) was used as a control.8
Cell culture
MA10 and R2C cells were cultured in Dulbecco's Modified Eagle's Medium (DMEM) supplemented with 15% horse serum (HS), 5% fetal bovine serum (FBS), and 100 U/mL penicillin and streptomycin. The cultures were maintained at 37℃ in a humidified atmosphere of 5% CO2. R2C cells were cultured in DMEM with 15% FBS and 5% HS, and infected with β-gal adenovirus (5 pfu) or TM FOXO3 virus (1 or 5 pfu) for 24 hr. The cultures were then maintained in serum free media for 48 hr. Testosterone levels in the medium were measured by radioimmunoassay (Diagnostic Systems Laboratories Inc., Webster, TX, USA), and cells were collected for a Western blot. R2C cells were plated and maintained in serum-free medium for 24 hr before treatment with 10 IU IVF-C (human chromic gonadotropin, LG Life Science, Seoul, Korea) or vehicle. To block the PI3K pathway, 20 µM of LY293002 were applied for 2 hr and then 10 IU of IVF-C were added.
Transfection and luciferase assay
One day before transfection, 293FT cells were transferred to 12-well plates. The mStAR reporter and expression plasmids were transfected at the indicated concentration for 6 hr with Lipofectamine 2000 (Invitrogen, Urlington, ON, Canada). After a set time, cell were lysed in 100 uL reporter lysis buffer (Promega Corp, San Luis Obispo, CA, USA) and assayed for luciferase activity using a lumat LB 9507 luminometer (EG&G, Berthold, Germany).
Immunofluorescence
For immunohistochemistry, 4 µm sections were cut using a Jung microtome (Leica, Heerbrugg, Switzerland). Sections were deparaffinized in xylene and hydrated in graded ethanol, followed by antigen retrieval in sodium citrate buffer. Sections were blocked in normal serum (5%) for 60 minutes and incubated with primary antibody at 4℃ overnight. After washing with Tris-buffered saline+0.05% Tween-20, Texas-red and fluorescein isothiocyanate-labeled donkey secondary antibodies (1:200, Jackson Immuno Research Lab) were added, and sections were incubated for 2 hr at room temperature. The slides were washed again and mounted with Vectashield medium (Vector Laboratories, Burlingame, CA, USA). A Zeiss Axioskop (Thornwood, NY, USA) was used to visualize the cells. Primary antibodies included rabbit polyclonal Foxo1 (1:500, SC-67140; Santa Cruz, CA, USA), Foxo3 (1:500, F2178; Sigma, MO, USA), and Foxo4 (1:200, A8975, Sigma) with goat anti-GATA-4 (1:200, Santa Cruz, CA, USA).
Western blot analysis
Cells were lysed in radioimmunoprecipitation assay buffer (20 mM Tris pH 7.5, 2 mM ethylenediaminetetraacetic acid, 150 mM NaCl, 0.5% phosphatase inhibitor, 0.5% protease inhibitors). Fifty grams of protein was separated on 10% sodium dodecyl sulfate gels, followed by semi-dry Western blotting. Subsequently, membranes were blocked with 5% bovine serum albumin (BSA) in TBST (50 mM Tris-HCl, pH 7.5 and 150 mM NaCl containing 0.05% Tween 20).19 The blots were probed overnight with Foxo3 (1:1000, Sigma F-2178), AKT (1:1000, Cell Signaling 4691), pAKT (1:1000, Cell Signaling 4060), and phospho-Foxo3 (1:1000, Cell Signaling 2599) in 5% BSA containing TBST (10 mM Tris pH 8.0, 150 mM NaCl, 0.05% Tween-20). After incubating with an anti-rabbit secondary antibody coupled with horseradish peroxidase, the immunocomplexes were visualized with enhanced chemiluminescence.
Statistical analysis
Quantitative data are expressed as mean standard deviation. Student's t-test (two-tails) was performed using SPSS 10.0 software (SPSS Inc., Chicago, IL, USA), and all p-values<0.05 were considered statistically significant. Data are presented as the mean±standard error of the mean.
RESULTS
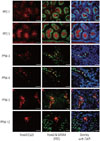
Foxo3 expression differed in Leydig cells according to age
We investigated the expression of Foxo3 using double-immunostaining with GATA4, which is expressed in Sertoli cells, steroidogenic Leydig cells, and other testicular somatic cells in the testes.20 As shown in Fig. 1, Foxo3 expression was abundant in spermatogonial cytoplasm during the pre-pubertal period but was undetected after puberty. Foxo3 expression in Leydig cells, however, began to appear at 3 weeks after birth and became more widespread. Most Foxo3 protein was detected in the nucleus at 3-4 weeks after birth. At 5 and 12 weeks, however, it was localized in both the nucleus and cytoplasm, which suggests active shuttling between these compartments. Foxo3 localization in Leydig cells was regulated by age and seemed to be related to its function.
hCG treatment increased Foxo3 phosphorylation in 30 min
As Foxo3 showed active shuttling in the Leydig cells around puberty, we hypothesized that LH hormone could be a major regulator of Foxo3 function, since Leydig cells are the primary site of LH hormone activity. Post-translational modification is a major mechanism that regulates the localization of Foxo proteins. Among various post-translational modifications, PI3K dependent phosphorylation of Foxo3 at Thr32, Ser253, and Ser315 residues is key regulator of excluding Foxo protein from the nucleus. We looked into the phosphorylation of Thr32 of Foxo3 by hCG in the R2C cells since LH and hCG are nearly identical at the molecular level. R2C cells treated with hCG exhibited increased levels of phosphorylated AKT at 30 min, which decreased thereafter. Foxo3 phosphorylation followed a similar pattern. This indicates that hCG stimulates the PI3K pathway to regulate the phosphorylation of Foxo3. To confirm this suspicion, we investigated the phosphorylation of Foxo3 after blocking the PI3K pathway by treatment with 20 uM of LY294002. Blocking the PI3K pathway decreased hCG-mediated phosphorylation of Foxo3 and AKT. This result confirmed that hCG/LH influences the phosphorylation of Foxo3 via the PI3K pathway, mediating whether Foxo3 stays in the cytoplasm or in the nucleus.
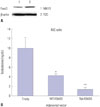
Testosterone levels decreased following FOXO3 virus infection
To elucidate the role of FOXO3 in Leydig cells, FOXO3 expression was confirmed in the Leydig cell lines R2C and MA10 (Fig. 3A). Both cell lines expressed FOXO3 protein, but levels thereof were higher in R2C cells. Because the main function of Leydig cells is to produce testosterone, hormone levels were measured in R2C cells over-expressing FOXO3. Adenovirus was used to express WT- and TM-FOXO3. TM-FOXO3 has alanine residues at each of the three AKT phosphorylation sites and is constitutively active in the nucleus.17 Over-expression of WT-FOXO3 decreased testosterone levels in R2C cells, which decreased further following TM-FOXO3 over-expression (Fig. 3B). This indicates that FOXO3 negatively regulates testosterone synthesis and is more active in the nucleus.
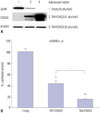
StAR protein levels following FOXO3 virus infection
To determine how FOXO3 decreases testosterone levels in R2C cells, we investigated StAR protein levels by TM-FOXO3. In testosterone production, cholesterol transportation to the inner membrane of mitochondria is a rate-limiting step. StAR is located in mitochondria and plays a key role in steroid hormone synthesis by transporting cholesterol from the outer mitochondrial membrane to the inner membrane and converting it to pregnelone.21 TM-FOXO3 expression decreased StAR protein levels in a dose-dependent manner (Fig. 4A). To investigate whether FOXO3 directly regulates StAR gene expression, a reporter vector system was used. A mouse StAR reporter vector containing 2730 bps of the StAR promoter sequence was constructed. Cotransfecting WT FOXO3 with the reporter vector decreased mStAR promoter activity, and TM FOXO3 further decreased the reported activity (Fig. 4B). This finding corresponds with those for StAR protein levels, reflecting that FOXO3 directly regulates StAR gene expression.
DISCUSSION
While the role of Foxo3 in the female reproductive system has been widely studied and clarified, little is known about the involvement of Foxo3 in male gonad development. In this study, the presence and location of Foxo3 transcription factors in mouse Leydig cells, the regulation of its location, and its role in testosterone production were identified. We observed that Foxo3 is mainly located within the testicles from birth to PPD 5. However, after PPW 3, Foxo3 was mainly located in Leydig cells, and its expression pattern depended on age. At PPW 3 and 4, most Foxo3 was detected in the nucleus. Later, it was detected in cytoplasm and the nucleus, indicating that Foxo3 localization is regulated in Leydig cells. Thus, Foxo3 location plays an important role in the activity of Leydig cells. The importance of Foxo protein in male reproductive system is also relfected by the fact that Foxo1 and Foxo4 are also detected in the testes (data not shown). Indeed, the importance of Foxo1 on spermatogonium has been emphasized in a previous study in which germ line specific Foxo1 knock out mice showed defects in proliferative expansion, and thus, testes were small by PPD 21 due to spermatogonial arrest and failure of meiotic initiation.14
Nevertheless, Foxo3 was detected in both the cytoplasm and nucleus at 5-12 weeks after birth in Leydig cells, showing active shuttling between the nucleus and the cytoplasm. Therefore, we hypothesized that puberty hormone may be involved in regulating gonadal development. Gonadotropins FSH and LH were expected to regulate Foxo3 localization because these gonadotropins are secreted by the pituitary and act on the ovaries and testes to initiate sexual maturation and to maintain a cyclic reproductive function.2223 Among gonadotropins, LH was a candidate factor for on Foxo3 localization in Leydig cells. In females, steroidogenesis is primarily controlled by LH in Theca cells.222425 LH stimulates Theca cells to produce testosterone and maintains progesterone production by inducing the genes involved in steroidogenesis: cytochrome P450 side-chain cleavage enzyme (CYP11A1), 3β-hydroxysteroid dehydrogenase, 17α-hydroxylase/C17-20 lyase cytochrome P450 (CYP17A1), and StAR protein.2627282930 In bovine Theca cells, Akt is constitutively expressed, but it is gradually phosphorylated through LH exposure.31 AKT signaling by LH does not only influence females, but also male hormone synthesis. In primary cultures of immature rat Leydig cells, LH/chorionic gonadotropin stimulated ERK1/2 and Akt phosphorylation.253233 This result was replicated in our study (Fig. 2A). hCG treatment not only increased pAKT levels, but also pFoxo3 levels in 30 min. Activated Akt signaling by LH/hCG can increase Foxo3 phosphorylation at Thr32, Ser253, and Ser315, causing Foxo3 to be transported from the nucleus to cytoplasm. This finding indicates that LH released from the pituitary regulates Foxo3 localization. As PI3K pathway activates AKT signaling pathway, we blocked the PI3K pathway to confirm whether hCG stimulates induction of the phosphorylation of Foxo3 via the PI3K pathway. Pre-treatment of LY294002 (PI3K pathway inhibitor) reduced hCG-mediated pAKT and pFoxo3 levels, indicating the PI3K pathway regulates the localization of Foxo3 (Fig. 2B).
To determine the function of Foxo3 in Leydig cells, we measured testosterone levels in a Leydig cell line of R2C cells, as the main function of Leydig cells is testosterone synthesis. Our study revealed that FOXO3 down-regulates testosterone levels in Leydig cells (Fig. 3B). Testosterone synthesis consists of a series of steroid precursor and enzyme syntheses. The ratelimiting step in steroid synthesis is catalyzed by StAR protein, which transports cholesterol from the outer mitochondrial membrane to the inner membrane.3435 StAR protein levels decreased dramatically in a dose-dependent manner in R2C cells infected with TM-FOXO3 (Fig. 4A). Next, we observed whether mStAR promoter activity is directly down-regulated by FOXO3. FOXO3 down-regulated mStAR promoter activity, which was more significant with TM-FOXO3 expression (Fig. 4B). This result correlates with the reduced testosterone levels caused by over-expression of WT and TM-FOXO3 in R2C cells (Fig. 3B). From those results, we concluded that Foxo3 plays an important role in testosterone hormone synthesis. However, there is no reported phenotype in Foxo3 KO male mice. It seems to be that Leydig cells not only express Foxo3 but also Foxo4 (data not shown). Foxo4 seems to have similar function in Leydig cells.
Foxo3 may be regulated by other signaling pathways, such as insulin like growth factor (IGF)-1 signaling. In the testes, IGF-1 reaches its maximum expression at PPW 4 when pubertal testosterone rises.36 Growth factor have been shown to activiate AKT, which leads to the phosphorylation of Foxo3 at its 14-3-3 binding site and determines Foxo3 relocalization to the cytoplasm.9 This result with our observations led us to hypothesize that during the first four weeks, IGF-1 phosphorylates Foxo3, trapping it in the cytoplasm through interactions with 14-3-3. Once IGF-1 has reached its peak expression and decreases, LH signaling-mediating Foxo3 may become involved in the regulation of gonadal development by regulating testosterone hormone.
In summary, this study investigated Foxo3 localization and its role in the testes. Foxo3 is expressed in the testes, but its expression sites differ according to age. Foxo3 expression in Leydig cells appears in the nucleus before puberty, and it shows active shuttling in between the nucleus and the cytoplasm after puberty. We confirmed the role of Foxo3 in Leydig cells as a negative regulator for testosterone hormone by reducing StAR gene expression. The fact that phosphorylated Foxo3 by hCG/LH suggests a possible mechanism that LH stimulates testosterone levels via PI3K-dependent inhibition of Foxo3 action in Leydig cells.

 XML Download
XML Download