

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Spinal cord injury (SCI) is often associated with permanent disability as a result of sensory or motor functional defects caused by neuronal death and demyelination after primary physical damage.1 Treatment with a single modality is unlikely to result in complete recovery. Therefore, combined treatment might be effective for improving therapeutic efficacy. In the field of gene therapy, therapeutic gene expression controlled by hypoxia-specific or tissue-specific gene expression systems offers a possibility for efficient and safe gene therapy.23456
In the stem cell therapy field, transplantation with neural stem cells (NSCs) showed therapeutic potential in animal models with incurable disorders, including SCI.78 Interestingly, it has been reported that stem cells can be used as a gene delivery system9 and that stem cell-mediated gene therapy results in a better therapeutic outcome and more stable therapeutic gene delivery than single therapy with stem cells or therapeutic genes alone.1011121314
Considering that most stem cell therapies for SCI have used NSCs or neural precursor cells, which have high neuronal differentiation potentials, we chose a neuronal cell type-inducible transgene expression system (NSE::) for transgene overexpression in NSCs. We assumed that a combined treatment strategy based on NSCs and an transgene expression system controlled by a neuron-specific enolase (NSE) promoter would increase therapeutic efficiency. Finally, we demonstrated that it is suitable with NSCs for the establishment of a gene and NSC therapy platform.
MATERIALS AND METHODS
Plasmid construction
To confirm whether the NSE was suitable with NSCs for this combined treatment strategy, both a neuronal cell-type-inducible luciferase expression system (NSE::Luci) and a general luciferase expression system (SV::Luci) were made by pGL3 luciferase reporter vectors. SV::Luci was purchased from Promega (Madison, WI, USA, Cat.no E1761). Neuron-specific enolase (NSE) promoter was provided by Prof. Lee at Hanyang University, Seoul, Korea. The NSE region was inserted between NheI and BglII sites of the pGL3-basic vector (Promega, Cat.no E1751). The construction designs of the plasmid are shown in Fig. 1.
Cell culture
We used mouse NSCs in this study. NSCs (CRL-2925, ATCC, Manassas, VA, USA) were cultured in dulbecco's modified eagle medium (DMEM)/F12 and supplemented with 10% fetal bovine serum (FBS; Thermo Scientific, Waltham, MA, USA) and 1% penicillin and streptomycin (Invitrogen, Carlsbad, CA, USA). Approximately 4×105 cells were plated in six-well culture dishes (4×104 cells/cm2) and incubated at 37℃ in a humidified atmosphere containing 5% CO2.
For the other experiment, we used the human embryonic kidney 293 cell (293FT) provided from Prof. Kang at Yonsei University. Cells were cultured in DMEM supplemented with 10% FBS, 1% penicillin and streptomycin, 1X non-essential amino acid (NEAA; Invitrogen), and 1X sodium pyruvate (Invitrogen). Approximately 6×105 cells were plated in six-well culture dishes (6×104 cells/cm2) and incubated at 37℃ in a humidified atmosphere containing 5% CO2.
Gene transfection
Transfection was performed using Lipofectamine 2000 (Invitrogen) as previously described.9 Briefly, NSE::Luci, SV::Luci, or SV::DsRed (Control) plasmid (each 4 µg) were individually mixed with serum-free and antibiotic-free DMEM/F12 medium. Lipofectamine (10 µL) also was mixed with serum-free and antibiotic-free DMEM/F12 medium. Plasmid DNA solution was mixed with the lipofectamine-DMEM mixture and then incubated for 20 min at room temperature (RT). The DNA-lipofectamine mixture was added to NSCs or 293FT cells, and the cells were incubated at 37℃ in a humidified atmosphere containing 5% CO2.
Neural differentiation
For neural differentiation, NSCs were cultured in presence of retinoic acid (RA; Sigma, St. Louis, MO, USA) for 7 days. RA was dissolved in dimethyl sulfoxide (DMSO) (working solution, 10 mM) and diluted with culture media (final concentration, 0.01 µM, 0.1 µM, and 1 µM). Briefly, NSCs were seeded on poly-l-lysine (2 µg/cm2, Sigma)-coated glass coverslips at a density of 2×104 cells/cm2. After 24-h incubation, the medium was replaced with a complete medium with or without RA and then changed every 2 days. Neural differentiation was carried out for 7 days.
Cell transplantation
All protocols were approved by the Animal Care and Use Committee of the Medical Research Institute at Yonsei University College of Medicine. All experiments were conducted in accordance with international guidelines on the ethical use of animals, and the number of animals used was minimized. For anesthesia, ICR mice (male, 6 weeks, 30 g, OrientBio, Seongnam, Gyeonggi-do, Korea) were intraperitoneally injected with ketamine (100 mg/kg, Yuhan, Seoul, Korea) and rompun (10 mg/kg, Bayer Korea, Seoul, Korea). Laminectomy was performed at thoracic level 11. SCI was carried out using self-closing forceps (compression injury for 10 sec). Animals were divided into two groups, those treated with SV::Luci-NSCs and those treated with NSE::Luci-NSCs. NSCs (5×105 cells/2 µL) were directly injected into the spinal cord using a microinjection pump (injection rate, 1 µL/min). After transplantation, the exposure site was closed using suture wound clips. Cyclosporin A (10 mg/kg, Chong Kun Dang, Seoul, Korea) and cefazolin (20 mg/kg, Yuhan) were intraperitoneally injected daily.
In Vivo Imaging System (IVIS)
The In Vivo Imaging System (IVIS) Spectrum (Xenogen, Alameda, CA, USA) was used to detect luciferase expression. For in vivo imaging, mice were anesthetized with ketamine (100 mg/kg) and rompun (10 mg/kg), and then the substrate D-luciferin (150 mg/kg) was injected intraperitoneally. Twenty minutes after injection, bioluminescence images were captured for 10 min. Regions of interest (ROIs) were analyzed, and total quantification of bioluminescence was quantified using Living Image® (Xenogen) software. For in vitro imaging, the substrate D-luciferin (150 µg/mL) was added to the SV::DsRed-NSCs (Control), SV::Luci-NSCs, or NSE::Luci-NSCs in medium, followed by mounting and bioluminescence detection for 2-3 min.
Luciferase assays
A luciferase assay was performed 24-48 hrs after gene transfection or cell transplantation, and each sample (cells or tissues) were lysed with Pro-prep™ (INTRON Biotechnology, Gyeonggi, Korea). The concentration of protein in lysates was measured using a bicinchoninic acid protein assay kit (Pierce, Iselin, NJ, USA), following the manufacturer's protocol. Luciferase expression was measured using a luminometer for 10 s. The level of luciferase expression was expressed in relative light units (RLU) per mg of protein.
Immunocytochemistry
Differentiated cells were fixed in 4% paraformaldehyde, pH 7.4 (Merck, Darmstadt, Germany), for 10-15 min at RT. Samples were washed with phosphate buffered saline (PBS, Thermo) three times. Blocking was performed with 10% normal donkey serum in PBS containing 0.3% Triton X-100 for 60 min at RT. Primary antibodies against beta-III-Tubulin (TUJ1), microtubule-associated protein 2 (MAP2), neural cell adhesion molecule (NCAM), and neurofilament (NF) were incubated overnight at 4℃. Samples were washed three times with PBS, and secondary antibodies were added and incubated for 1 h at RT. Samples were washed three times with PBS, and 4',6'-diamino-2-phenylinodole (DAPI; Vector Labs, Burlingame, CA, USA) was added. Samples were covered with glass coverslips. A confocal laser scanning microscope (LSM 700, Zeiss, Oberkochen, Germany) was used for analysis.
Statistical analysis
Statistical analysis was performed using Prism 6 (Graphpad, San Diego, CA, USA). All data were evaluated using Student's t-test and a one-way analysis of variance (ANOVA). All data are expressed as the mean±standard error of the mean. All p-values<0.05 were considered statistically significant.
RESULTS
Expression pattern of neural stem cells transfected with a neuronal cell-type-inducible luciferase overexpression system (NSE::Luci-NSCs)
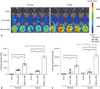
NSE::Luci was constructed using NSE promoter and pGL3-basic plasmid (Fig. 1). To confirm whether this neuronal cell-type-inducible gene overexpression system is suitable for NSCs, we investigated the expression level of luciferase in NSCs transfected with SV::DsRed (Control), SV::Luci plasmid, and NSE::Luci plasmid. Based on the results of IVIS and the luciferase assay, we confirmed that luciferase expression significantly increased in NSE::Luci-NSCs, compared with SV::Luci-NSCs (Fig. 2). These results indicate that the neuronal cell type-inducible transgene overexpression system is suitable for NSCs. To confirm whether the transgene overexpression pattern can be sustained after hypoxic injury (mimicking tissue ischemia after SCI), we investigated the luciferase expression level of Control, SV::Luci-NSCs, and NSE::Luci-NSCs after hypoxic injury for 24 h. We confirmed that the luciferase expression level significantly increased in NSE::Luci-NSCs, compared with that in SV::Luci-NSCs (Fig. 2). These results indicate that combined therapy with NSCs and a neuronal cell type-inducible gene overexpression system can be applied to spinal cord injuries that cause tissue ischemia.
To confirm whether the NSE was selectively working in neuronal lineage cells, we examined the luciferase expression level in 293FT cells, non-neuronal cell, transfected with SV::DsRed (Control), SV::Luci, or NSE::Luci plasmid. We confirmed that the luciferase expression level in 293FT transfected with NSE::Luci plasmid was significantly lower than 293FT transfected with SV::Luci plasmid (Fig. 3). This result means that the efficacy of the neuronal cell type-inducible gene expression system depends on neuronal lineage cell type.
Transgene expression pattern in differentiated neurons
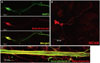
To confirm the differentiation potency of NSCs, neural differentiation was induced by RA treatment. Differentiated cells cultured in the presence of 1 µM RA for 7 d were stained for neuronal specific markers, such as TUJ1, MAP2, NCAM, and NF. We confirmed that differentiated cells were positive for all neuronal markers (Fig. 4), meaning that NSCs used in this experiment were efficiently differentiated into neurons.
To confirm whether the luciferase expression pattern of the NSE::Luci-NSCs can be increased after neuronal differentiation (1 µM RA), compared with NSC stage (0 µM RA), we investigated the luciferase expression level at the stage of NSCs and differentiated neurons after transfection with NSE::Luci or SV::Luci plasmids. We confirmed that the luciferase expression level of NSCs transfected with NSE::Luci plasmid was much higher after neuronal differentiation (Fig. 5A). At the neuronal differentiation condition (1 µM RA), the luciferase expression level of NSE::Luci-NSCs was significantly higher than that of SV::Luci-NSCs (Fig. 5B).
These results mean that the transgene expression by NSE promoter was induced by neuronal differentiation. Thus, these findings indicate that the neuronal cell-type-inducible transgene overexpression system appropriates with NSCs, which have a neuronal differentiation potency for combined treatment strategies.
Transgene expression pattern in vivo
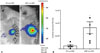
To confirm whether NSE::Luci-NSCs show a consistent high expression pattern in vivo, both SV::Luci-NSCs and NSE::Luci-NSCs were transplanted into the spinal cord, and an IVIS analysis and luciferase assay was carried out 24-48 h after transplantation. Similar to our results in vitro, the luciferase expression level was significantly higher in NSE::Luci-NSCs than in SV::Luci-NSCs (Fig. 6). These results indicate that a combined treatment strategy based on a neuronal cell type-inducible transgene expression system and NSCs can be applied to SCI.
DISCUSSION
As one of the pathologies that appear after SCI, an ischemic environment in the spinal cord is caused by the destruction of blood vessels; this ischemic condition lasts from several days to several weeks. The survival rate of stem cells transplanted into the ischemic environment would normally rapidly decline. For the protection of transplanted stem cells and the injured spinal cord, we designed a gene and stem cell therapy platform based on NSCs and a neuronal cell type-inducible gene expression system.
Gene overexpression by NSE promoter is already known to work in mature neurons. However, we confirmed that the luciferase expression level of NSE::Luci-NSCs was significantly higher than the luciferase expression level of SV::Luci-NSCs, and the luciferase expression level of 293FT cells (used as a mean non-neuronal cell type) transfected with NSE::Luci plasmid was significantly lower than that of 293FT cells transfected with SV::Luci plasmid. This result means that transgene overexpression by NSE promoter is already working at the NSC stage. A previous study investigated the difference of gene overexpression based on NSE promoter between neurons and non-neuronal cells, such as HEK293 and HeLa cells; however, the study did not examine the difference of gene overexpression at the NSC stage.4 Thus, we carefully suggest that neuron-specific promoter may function extensively in neuronal lineage cells including NSCs.
Additionally, we confirmed that NSE::Luci-NSCs showed a higher luciferase expression level than the SV::Luci-NSCs under normoxia and hypoxia, and the luciferase expression level of NSE::Luci-NSCs was much higher after differentiation into neurons. All of these features can be used advantageously, if the luciferase used in this study were to be replaced by other therapeutic genes.
In our previous study, we confirmed that a NSCs transfected with a hypoxia-inducible gene expression system should be maintained for at least 1 day under hypoxic conditions for transgene overexpression.9 However, the gene expression level of NSCs transfected with a neuronal cell type-inducible gene overexpression system was sufficiently greater in normoxia, namely before transplantation (saving time, as there is no need to wait 1 day). Thus, the high amount of secreted therapeutic protein within 1 day may be useful for improving the survival of transplanted stem cells, as well as protecting the injured spinal cord from exposure to an ischemic environment. Subsequently, the high amount of secreted therapeutic protein from transplanted NSCs would be sustained under the ischemic environment and would also increase further neuronal differentiation.
Thus, these findings suggest that a neuronal cell type-inducible gene expression system is suitable for use with NSCs for expressing a high amount of therapeutic genes, and such gene and NSC therapy based on the present systems described here appear to be very promising as potential treatments for SCI.
In this study, we proposed a combined strategy using a neuronal cell type-inducible transgene overexpression system and NSCs for SCI treatment. We confirmed that the luciferase gene expression level of NSE::Luci-NSCs was significantly higher than that of SV::Luci-NSCs. This pattern was sustained after hypoxic injury and in vivo. Specifically, the luciferase gene expression level observed at the NSC stage was much greater after differentiation into neurons. We suggest that a neuronal cell type-inducible transgene overexpression system is suitable for NSCs; furthermore, a combined treatment strategy based on a neuronal cell type-inducible therapeutic gene overexpression system and NSCs may be a promising tool for the treatment of SCI.



 XML Download
XML Download