

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
House dust mites (HDMs) are one of the most common aeroallergens worldwide and a crucial trigger for allergic responses (asthma, rhinitis and atopic dermatitis).1,2 Exposure to HDMs in sensitized patients is associated with worse lung function, greater medication requirements, and the development of chronic asthma.3,4,5 Both D. pteronyssinus and D. farinae are two of the most predominant species, the major allergens of which are group 1 allergens: Der p 1 and Der f 1. These allergens have diverse biological functions (e.g., cysteine proteases) that contribute to allergic sensitization and share amino acid sequences with high homology.6,7,8
IgY is the principal known antibody in amphibians, reptiles, and birds.9 In hens, IgY is transported from the blood to the egg yolk, where it accumulates and provides offspring humoral immunity against avian pathogens prior to full maturation of their own immune system.10,11,12 IgY antibody concentrations range from 50 to 100 mg per egg yolk.12 Although chicken IgY shares a common ancestor with both mammalian IgG and IgE, the egg yolk antibodies neither activate the mammalian complement system, nor interact with rheumatoid factors or bacterial and human Fc receptors that could mediate an inflammatory response.11,12,13 In recent studies, chicken egg yolk has been recognized as an alternative antibody source. Immunization with egg yolk immunoglobulin (IgY) has shown diagnostic and therapeutic value against fungi, bacteria and virus causing various diseases.14,15,16,17 For example, passive immunization with egg yolk IgY through intramuscular injection, oral administration, and immersion was shown to have therapeutic value in the treatment of infectious disease evoked by rotavirus, parvovirus, E. coli, S. typhimurium, S. mutans, H. pylori, and Candida albicans.16
In this study, we aimed to produce IgYs, which are specific to D. pteronyssinus and D. farinae, from hens and to examine immunoreactivities for further diagnostic use of HDMs specific IgYs.
MATERIALS AND METHODS
HDM extracts
HDM (D. pteronyssinus or D. farinae) was pulverized in liquid nitrogen. The fatted samples were then placed in 1:1 ethyl ether/ethyl acetate and extracted with slow stirring at 4℃ for 2 days in phosphate buffered saline (PBS) (pH 7.4). The extract was then centrifuged at 9000×g for 30 minutes at 4℃, and the supernatant was dialyzed overnight and lyophilized. The extract was diluted in 0.01 M potassium phosphate buffer (pH 7.2) containing 0.1 M NaCl and mixed with Freund's complete adjuvant (FCA, Sigma, MO, USA) at a 1:1 ratio. The protein concentration of HDM was determined by Bradford protein assay and the protease activity was measured by QuantiCleave protease assay kit (Pierce, Rockford, IL, USA). The protease activity of D. pteronyssinus and D. farinae were 5 U/mL and 4.78 U/mL (per protein extract 50 ug/mL), respectively.
Immunization of hens
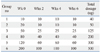
For the induction of HDM-specific antibodies in egg yolks, 22-week-old White Leghorns were immunized intramuscularly (specifically, in the breast tissue) with a mixture of HDM protein extract and adjuvant under conventional animal room conditions that were mite-free. Booster inoculations consisting of half the initial volume were administered to hens at week 2 and repeated in the same manner every 2 weeks (Group 2-6) (Table 1). The hens of the control group received inoculations in the same manner via a sterile saline solution (PBS). The eggs were harvested from start to finish of the experiment and stored in a refrigerator at 4℃.
Isolation and purification of IgY
Eggs were individually cracked, and the egg white was separated from the yolk. Residual egg white was carefully wiped off using a gauze sheet so as not to break the yolk. The yolk sac was ruptured with a needle and the contents were dripped into a tared beaker. After discarding the yolk sac, the weight of the separated yolk was measured. IgY from the egg yolk was purified using an EGGstract IgY purification kit (Promega, WI, USA). The method used was partly modified from that of Polson, et al.18 using polyethylene glycol and ammonium sulfate precipitation. Then, the precipitated IgYs were dissolved in PBS and stored at -20℃ until analysis.
Specific activity of IgY
Each HDM antigen (50 ug/mL per well) was coated previously onto an enzyme-linked immunosorbent assay (ELISA) plate for 16 hours, and diluted (1:500-1:2000) IgY samples and standards were then applied and incubated for 1 hour. Standard IgYs were produced by passing through an affinity column conjugated purified HDM. After washing 3 times, the attached IgYs were reacted with anti-IgY-alkaline phophatase conjugate for 1 hour for colorization (Promega, WI, USA). The absorbance was measured at 405 nm to determine the quantities of IgY specific to HDMs. The unit of IgY specific to HDM (ug/mg) was defined as the value of IgY specific to HDMs divided by the total protein concentration of IgY calculated in every egg, given that the amount of IgY in yolk is independent of egg weight.
Western blotting analysis
Sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) was performed using reducing and denaturing conditions. Protein extracts from HDMs were loaded onto 12.5% gels and stained with Coomassie blue to visualize constitutive protein bands or for Western blotting analysis. The separated proteins were electro-transferred onto nitrocellulose membranes and blocked with 5% skim milk. These strips were incubated with purified IgY diluted into different ratios (v/v) and then reacted with anti-chicken IgY alkaline phosphate conjugate (Promega, WI, USA) after additional washing. Bands were made visible on the membrane using 5-bromo-4-chloro-3-indolyl-phosphate/nitro blue tetrazolium (Promega, WI, USA) color development substrates.
RESULTS
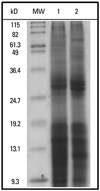
Extracted proteins of D. pteronyssinus and D. farinae were analyzed by SDS-PAGE. Protein bands less than 36.4 kD were strongly stained with Coomassie blue in HDMs, including Der p 1 (25 kD) and Der f 1 (36 kD), major allergens of HDMs with known proteolytic activity and maximum immunogenicity (Fig. 1).8
The HDM extracts were intramuscularly inoculated in hens 3 times every 2 weeks (Table 1), and eggs were harvested every day. IgYs were purified from egg, and IgYs specific to HDMs were detected. As shown in Fig. 2A, IgY specific to D. pteronyssinus started to increase at day 6 (89.0 ug/mg) after the first immunization at day 1 and rapidly increased from days 14 to 16 (92.8-760.2 ug/mg) when the second injection was carried out at day 14. These results suggest that the first inoculation caused a surge in specific IgYs against D. pteronyssinus. After reaching a peak at day 16 (760.2 ug/mg), however, these IgYs gradually decreased by day 29 (435.0 ug/mg), despite a third immunization at day 28. From day 30 onward, IgY specific to D. pteronyssinus was not significantly distinguished from the control. With the fourth injection, the last peak simultaneously appeared at day 42 (278.3 ug/mg) and immediately dropped toward control levels since day 43 (56.3 ug/mg), indicating the fourth injection did not affect specific IgY productivity. On the other hand, IgYs specific to D. farinae were continuously induced until day 47 (661.3 ug/mg), and an increase in these specific IgYs was detected after each immunization: at day 2 (134.9 ug/mg), 22 (425.9 ug/mg), 39 (517.1 ug/mg) and 47 (661.3 ug/mg) (Fig. 2B). These results suggest that D. farinae was more potent in producing and maintaining high amounts of specific IgY in the yolk after each injection than D. pteronyssinus.
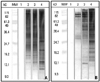
Furthermore, we attempted to visually analyze the immunoreactivities of purified IgYs specifically against HDMs by Western blotting (Fig. 3). The immunoreactivity of HDM protein bands for specific IgYs was dependent on the dilution ratio of IgY, and IgYs from control hens did not react to any HDM protein bands. The IgYs used for Western blotting were chosen based on the highest value of IgY specific to D. pteronyssinus or D. farinae.
DISCUSSION
In this study, we produced IgY specific to HDM protein from eggs harvested after immunization, and confirmed that the purified specific IgY responded dose-dependently to D. pteronyssinus or D. farinae protein. For six weeks, including four successive sensitizations, antibody titers differed between D. pteronyssinus and D. farinae immunization groups; the anti-D. farinae group continued to produce stable results after an additional booster compared with the anti-D. pteronyssinus group (134.9-661.3 ug/mg) (Fig. 2A). Even though the productivity of anti-D. pteronyssinus group was relatively malleable, however, antibody titers remained very high in the anti-D. pteronyssinus group (40.9-760.2 ug/mg) (Fig. 2B). These findings indicate that both D. pteronyssinus and D. farinae potently induce specific IgYs, suggesting that at least a second immunization should be carried out to induce a maximal antibody level response.
In cases of D. pteronyssinus, over 30 different proteins have been found to induce IgE antibody in HDM allergic patients; 21 of these allergens demonstrated cross-reactivity with allergens from other mite species.19 Although they show immunologic cross-reactivity, D. pteronyssinus and D. farinae typically have 15-20% amino acid sequence disparity and also have unique epitopes.20 Just as anti-inflammatory medicines and immunosuppressive agents may be used to combat inflammation, allergic symptoms to HDM may be relieved by specific immunotherapies.21 Xie reported that oral administration of D. farinae extract significantly reduced goblet cell hyperplasia and eosinophilia in a D. farinae-sensitized allergic rat model.22 However, even though recent studies have emphasized the requirement for specific immunotherapy as an allergen-specific approach, they have not addressed the difficulties encountered in allergen standardization and the risk of inducing life-threatening anaphylactic side effects. Thus, efforts to develop specific antibodies against various HDM allergens absolutely requires testing proposed risk levels and assessing the corresponding airborne allergen levels, which constitute risk factors.23 Furthermore, these antibodies will also be useful in standardizing allergen extracts used for diagnosis and immunotherapy, as well as for comparing the efficacy of allergen avoidance regimes. In addition, this will make it easier to provide information on the relationship between mite allergen levels and the prevalence of allergic disease.24 For these antibodies, the use of chicken IgY instead of mammalian antibodies has numerous advantages, including increased production of antibody levels, decreased animal use, and no need for painful blood collections. The utility of IgY without a loss of specificity and sensitivity has been demonstrated in many immunological assays, and maintaining chickens was more cost-effective compared to other mammalians.12,25 Moreover, chickens from inbred strains are obtainable, thus minimizing genetic variation in antibody responses. Yolk antibodies show increased deposition in the egg during the process of transmission from the maternal line to offspring.26,27,28
In conclusion, we immunized hens with HDMs and confirmed a significant increase in the level of IgYs specific to HDMs. A number of immunoreactive proteins involving major HDM allergens of approximately 9.3 to 115 kD were also detected. Though further work is required to confirm the immunoreactivity of specific IgYs in patient sera sensitized to HDMs, our results suggest that chicken egg yolk antibodies have considerable potential as an alternative antibody source. This could be a major source of research opportunities, including diagnostic analysis and immunotherapy development in the field of allergic disease.

 XML Download
XML Download