

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
The prefrontal cortex (PFC) is involved in numerous cognitive functions, such as working memory, attention, arousal, decision making, execution, behavior inhibition, and motivation.1,2 Each subregion of the PFC has a specific role in cognition. The main functions of the four subregions of the PFC are as follows: dorsolateral prefrontal cortex (DLPFC)-working memory, ventrolateral prefrontal cortex (VLPFC)-deliberation of decision making and goal-directed behavior, the orbitofrontal cortex (OFC)-emotional control and inhibitory control of behavior, and the medial PFC-motivation and initiation of activity.1,2,3,4,5 These PFCs receive copious afferent fibers from the mediodorsal nucleus (MD) of the thalamus via anterior thalamic radiation.6
Injury of the PFC can cause frontal syndrome, which includes cognitive and emotional dysfunctions.1 On the other hand, patients with a lesion in the MD of the thalamus can exhibit clinical manifestations similar to those associated with a PFC injury, possibly because of thalamocortical connections between the MD of the thalamus and the PFC.7,8,9 Furthermore, frontal network syndrome has also been referred to patients with a lesion within the thalamocortical connections between the MD of the thalamus and the PFC.1 Furthermore, these connections are also known to be involved in many psychiatric diseases, such as, schizophrenia, addiction, depression, and bipolar disorder.2,10 Therefore, the elucidation of these thalamocortical connections would undoubtedly be useful in the clinical field in terms of neurorehabilitation and psychiatry. Many animal studies have demonstrated these thalamocortical connections,6,11,12,13,14,15,16,17 however, detailed neural pathways of these connections in human brain have not yet been identified.18,19
The recent development of diffusion tensor tractography (DTT), which was derived from diffusion tensor imaging (DTI), allows visualization and localization of neural tracts at the subcortical level in three dimensions. DTT enables the state of thalamic radiations and thalamus-related neural tracts to be assessed in human brain.20,21,22,23,24,25 Nevertheless, little is known about the thalamocortical connections between the MD of thalamus and the PFC in human brain.18,19
The present study was undertaken to identify and investigate the anatomical characteristics of thalamocortical connections between the MD and the PFC in human brain using DTT.
MATERIALS AND METHODS
Subjects
Thirty-two normal healthy subjects (19 males and 13 females of mean age 36.94±11.51 years; range 20 to 62) with no history of neurologic disease were recruited for this study. All participants provided written consent, and the study was approved by the Institutional Review Board of our university hospital.
Diffusion tensor imaging
DTI data were acquired using a 6-channel head coil on a 1.5 T Philips Gyroscan Intera (Philips, Best, the Netherlands) and single-shot echo-planar imaging. For each of the 32 non-collinear diffusion sensitizing gradients, we acquired 67 contiguous slices parallel to the anterior commissure-posterior commissure line. Imaging parameters were as follows: acquisition matrix=96×96; reconstructed matrix=192×192; field of view=240×240 mm2; TR=10726 ms; TE=76 ms; parallel imaging reduction factor (SENSE factor)=2; EPI factor=49; b=1000 s/mm2; NEX=1; and a slice thickness of 2.5 mm with no gap.
Probabilistic fiber tracking
Diffusion-weighted imaging data were analyzed using software from the Oxford Centre for Functional Magnetic Resonance Imaging of the Brain (FMRIB) Software Library (FSL v5.0; www.fmrib.ox.ac.uk/fsl). Affine multi-scale two-dimensional registration was used for correction of head motion and eddy current-induced image distortion. Fiber tracking was performed using a probabilistic tractography method based on a multiple tensor model, using tractography routines implemented in FMRIB Diffusion (5000 streamline samples, 0.5 mm step lengths, curvature thresholds=0.2).26
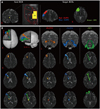
In each case, a seed region of interest (ROI) was placed at the MD of the thalamus on a coronal image of b0 image (no diffusion weighting applied). MD location was defined as previously described.19,27 For thalamocortical connections from the MD to the DLPFC and VLPFC, we defined the DLPFC as Brodmann areas (BAs) 8, 9, and 46, and the VLPFC as BAs 44, 45, and 47, and manually drew target ROIs on the DLPFC and VLPFC on the coronal images, respectively (Fig. 1).19,28 For thalamocortical connection from the MD to the OFC, we defined the OFC as BAs 47/12, 10, 11, and 13, and manually drew the target ROI on the OFC on an axial image.19,29 Thalamocortical connections between the MD and the PFC were determined by selecting fibers passing through seed and target ROIs. Of 5000 samples generated from the seed voxel, contact results were visualized at a threshold minimum of 1 streamline through each voxel for analysis. Fractional anisotropy (FA), mean diffusivity, and tract volume in the three thalamocortical connections to each PFC were then measured.
Statistical analysis
SPSS software (v.15.0; SPSS, Chicago, IL, USA) was used for the data analysis. One-way analysis of variance (ANOVA) with Tukey's post-hoc test was used to determine significances of difference in DTI parameters between reconstructed thalamocortical connections. Statistical significance was accepted for p values of <0.05.
RESULTS
The three reconstructed thalamocortical connections between the thalamic MD and the PFC are shown in Fig. 1. These thalamocortical connections were arranged in subcortical white matter in the following order; DLPFC, VLPFC, and OFC. Although some individual variations were observed, the main courses of these three thalamocortical connections were as follows; thalamocortical connections to the DLPFC ascended through the anterior thalamic radiation via the anterior limb and genu of the upper internal capsule, the corona radiata around the anterior horn and the anterior centrum semiovale, and then terminated at the DLPFC. Thalamocortical connections to the VLPFC also ascended through the anterior thalamic radiation via the anterior limb and genu of the upper internal capsule and the corona radiata around the anterior horn, and then terminated at the VLPFC. Thalamocortical connections to the OFC passed horizontally through the anterior thalamic radiation via the anterior limb and genu of the lower internal capsule and terminated at the OFC (Fig. 1). At the centrum semiovale level, thalamocortical connections to the DLPFC were located anterior to the genu of the corpus callosum. By contrast, at the corona radiata level, thalamocortical connections to the VLPFC were located around the frontal horn of the lateral ventricle anterior to the thalamocortical connection to the DLPFC. At the internal capsule, the thalamocortical connections to the OFC were located at the lower internal capsule below the thalamocortical connections to the DLPFC and VLPFC, which were usually located at the upper internal capsule.
Mean values for FA, MD, and tract volume of DLPFC were 0.33±0.03, 0.85±0.06, and 930.03±435.70, respectively, and those of VLPFC were 0.33±0.02, 0.85±0.05, and 1379.81±546.09, respectively. In the OFC, average FA, mean diffusivity, and tract volume were 0.33±0.02, 0.83±0.05, and 2139.00±802.62, respectively. In terms of FA and mean diffusivity, no significant differences were observed between the DLPFC, VLPFC, and OFC (p>0.05) (Table 1). Furthermore, no significant difference was observed between average DLPFC and VLPFC tract volume (p>0.05), but average OFC tract volume was significantly greater than DLPFC and VLPFC tract volume (p<0.05) (Table 1).
DISCUSSION
In the current study, thalamocortical connections between thalamic MD and DLPFC, VLPFC, and OFC in human brain were reconstructed using DTT. In addition, we characterized thalamocortical neural tracts and their courses between thalamic MD and three PFCs. Thalamocortical connections to DLPFC were located at the centrum semiovale anterior to the genu of the corpus callosum, and thalamocortical connections to VLPFC were located around the frontal horn of the lateral ventricle anterior to the thalamocortical connections to DLPFC at the anterior corona radiata. At the internal capsule level, thalamocortical connections to OFC were located at the lower anterior limb and genu of the internal capsule below the thalamocortical connections to the DLPFC and VLPFC, which were mainly located at the upper anterior limb and genu of the internal capsule. Thus, these thalamocortical connections had different pathways at the centrum semiovale, corona radiata, and internal capsule levels in the order of DLPFC, VLPFC, and OFC, that is, from an upper to a lower level.
Thalamocortical connections between thalamic MD and DLPFC, VLPFC, and OFC have been described in many animal studies, the majority of which focused on connectivity between thalamic nuclei and PFC.11,12,13,14,15,16,17 Some DTI studies on human brain have been reported to date.18,19 Using DTT, Klein, et al.19 found a high probability of connection between parvicellular MD and lateral PFC, between the fibrous part of MD and lateral OFC, and between the caudodorsal part of MD and the medial PFC in human brain. In 2011, Eckert, et al.18 reported a different connectivity of MD and of the centromedian-parafascicular complex of thalamus to PFC and subcortical regions, and that thalamic MD is more frequently connected to PFC than the centromedian-parafascicular complex of thalamus. Recently, using DTI, Kubota, et al.30 reconstructed entire thalamocirtocal pathway, thalamolateral and thalamomedial prefrontal pathway and thalamo-orbitofrontal pathway in schizophrenia patients, and found that schizophrenia patients showed decreased FA value in the thalamo-orbitofronal pathway compared with normal healthy subjects. We performed this study because these previous two DTI studies provided little information on the neural pathways of thalamocortical connections, which are clinically important because they are involved in frontal network syndrome as well as in many psychiatric diseases.1
In conclusion, we identified thalamocortical connections between MD of thalamus and three PFCs in human brain, using DTT. We believe that the methodology used and results of this study would be helpful for researchers in the neuroscience field and also to those involved in neurorehabilitation and psychiatry. In particular, the described courses of the three thalamocortical connections should be useful to clinicians treating frontal network syndrome. However, this study is limited by our inability to reconstruct thalamocortical connections to the medial PFC, and we consider that this topic is worthy of future study. In addition, further studies on clinical correlations and on the reliabilities and validities of the three thalamocortical connections to PFCs are needed.

 XML Download
XML Download