

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Rosacea is a common inflammatory skin disease, most prominent in the central area of the face, namely the cheeks, nose, chin, and forehead, which are densely populated with sebaceous glands.1 Fair-skinned individuals suffer disproportionately from this disease with its chronic relapsing course. The disease is characterized by primary and secondary features: primary features include flushing, persistent erythema, telangiectasia, as well as papules and pustules, while secondary features consist of burning or stinging, plaques, dry appearance, edema or phyma.2 The chronic inflammatory and vascular manifestations of rosacea point to a complex, multifactorial pathophysiology. Interestingly, recent findings suggest that an altered regulation of the innate immune response may be involved.3
Cathelicidins comprise a family of antimicrobial peptides (AMPs) expressed in leukocytes and in various epithelial tissues, where they participate in innate immune responses.4,5 Some cathelicidin peptides, including LL-37 expressed in humans, express both vasoactive and proinflammatory activities.6,7 Significantly higher levels of cathelicidin peptides may be detected in rosacea-affected skin, compared to healthy skin.8 Increased expression of the serine protease kallikrein 5 in the skin of patients with rosacea may promote proteolytic conversion of cathelicidin precursor into the active cathelicidin peptide LL-37.8,9 Abnormally increased expression of LL-37 may induce vascular endothelial growth factor (VEGF) in epidermal keratinocytes, which corresponds to the enhanced inflammation and vascular response seen in rosacea.9,10
Protease-activated receptor 2 (PAR-2), a PAR-family G-protein-coupled 7-transmembrane domain receptor, is activated by serine proteases such as kallikrein, and PAR-2 mediates inflammation in various tissues.11,12,13,14 PAR-2 signaling may also promote expression of the epithelial antimicrobial peptides human β-defensin-2 (hBD-2) and LL-37 as components of innate immunity.15,16 Although expression of the serine protease that activates PAR-2 is increased in rosacea, the regulation of PAR-2 and its role in rosacea remain unclear.
We hypothesized that PAR-2 expression increases in the skin of patients with rosacea and that direct activation of PAR-2 would increase production of cathelicidin LL-37, which may participate in rosacea. To elucidate the role of PAR-2 in the pathogenesis of rosacea, we tested for correlations between cathelicidin and PAR-2 expression in skin samples from patients with rosacea, as well as the induction of cathelicidin expression in cultured keratinocytes upon treatment with PAR-2 activating peptide (AP).
Go to : 
MATERIALS AND METHODS
Patient samples
The Institutional Review Board of Severance Hospital approved this study (No. 4-2012-0191). Data for the 40 patients diagnosed with rosacea at the Department of Dermatology, Yonsei University College of Medicine from 2005 to 2010 were reviewed. The diagnosis of rosacea was based on clinical photos and medical charts together with pathological findings. For comparison, 20 samples of facial skin diagnosed as no specific findings or milium without inflammation were selected for review.
Cell culture and reagents
Cells of the immortalized human keratinocyte cell line HaCaT were maintained in Dulbecco's Modified Eagle's medium (Welgene, Daegu, Korea) supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin at 37℃ in a humidified CO2 incubator (95% air, 5% CO2). PAR-2 AP, SLIGKV-NH2 (Ser-Leu-Ile-Gly-Lys-Val) (Peptron, Daejeon, Korea) was used to activate PAR-2. Reverse PAR-2 control peptide (CP) (Peptron) was used to determine the specificity of effect of PAR-2 activators. PAR-2 AP (100 µM) was added to HaCaT cells after replacing the culture medium with serum-free medium. In this time-dependent stimulation experiment, HaCaT cells were all plated at the same time, and PAR-2 was then added at the appropriate times thereafter. All HaCaT cells were simultaneously harvested 24 hours after plating.
Histology and immunohistochemistry
Hematoxylin-eosin stained slides and paraffin-embedded tissues from each case were retrieved from the surgical files of the Department of Pathology. Each tissue was serially sliced into four sections 3 to 4 µm thick for immunohistochemical analysis. Immunohistochemistry was performed using the polymer detection method (NovoLink Max Polymer Detection System, Leica Biosystems, Newcastle Ltd., Newcastle, UK).
Briefly, sections previously attached to silane-coated slides were dried at 60℃ for 1 hr. The sections were deparaffinized in xylene and rehydrated in graded alcohol and distilled water. For antigen retrieval, the sections were put into 10 mM citrate buffer (pH 6.0) and boiled in a microwave oven. After cooling at room temperature, sections were treated with a peroxidase block kit for 15 min, washed with Tris-buffered saline (TBS) for 5 min, and incubated with primary antibodies at room temperature for 1 hr.
The antibodies and dilutions used were as follows: PAR-2 (rabbit polyclonal anti-human, Santa Cruz Biotechnology, Santa Cruz, CA, USA, 1:200) and cathelicidin (rabbit polyclonal anti-human, Abcam, Cambridge, MA, USA, 1:200). Slides were washed with TBS three times, and then reacted with Post Primary (included in the kit) at room temperature for 30 min. After washing with TBS three times, slides were reacted with polymer for 30 min. Color was developed using the NovaRED substrate kit (VECTOR Laboratory, Burlingame, CA, USA). The reaction was stopped with distilled water while being observed under a microscope. Harris hematoxylin was used as a counterstain.
Slides were examined at 400× magnification. Two investigators independently reviewed slides without knowledge of the clinical data. The intensity of keratinocytes showing immunoreactivity to each marker in more than three high powered fields was assessed semi-quantitatively using the following grading system: 0=no staining, 1=weak staining, 2=moderate staining, and 3=strong staining.
Real-time RT-PCR analysis
Total RNA was extracted using the RNeasy mini-kit (Qiagen, Valencia, CA, USA) according to the manufacturer's instructions. One microgram of total RNA was used as the first strand for cDNA synthesis with Primescript™ (TaKaRa Bio Inc., Otsu, Japan). For all quantitative real-time RT-PCR analyses, amplification was performed in a Step One Real-Time PCR System (Applied Biosystems, Foster City, CA, USA) and visualized with a Power SYBR GREEN MASTER MIX (Applied Biosystems). The cycling conditions were as follows: 2 min at 50℃, 10 min at 95℃, and 40 cycles of 95℃ for 15 seconds, and 60℃ for 1 min. For each specific mRNA analysis, gene expression was normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) mRNA in the same sample. The primer sequences used were as follows: PAR-2 forward, 5'-GGCCCTGTGGGTCTTTCTTTT-3', and reverse, 5'-CAAGGGGAACCAGATGACAGA-3'; forward 5'-GGCTGGTGAAGCGGTGTAT-3', and reverse, 5'-TGGGTACAAGATTCCGCAAAAA-3' (cathelicidin); forward, 5'-CGCAGCTACTGCCATCCAAT-3', reverse, 5'-GTGAGGTTTGATCCGCATAATCT-3' (VEGF); forward, 5'-GCTCCTCCTGTTCGACAGTCA-3' and reverse, 5'-ACCTTCCATGGTGTCTGA-3' (GAPDH).
Relative quantification was performed using the ΔΔCt (threshold cycle) method, where ΔCt was (target Ct-GAPDH Ct), and ΔΔCt was (ΔCt sample-ΔCt untreated control). The quantity of the sample relative to control mRNA was 2-ΔΔCt. The mRNA expression was indicated with a Ct value and was corrected for GAPDH expression.
Western blotting
Whole cell extracts were prepared in RIPA buffer (TBS, 0.5% deoxycholate, 0.1% SDS, 1% Triton X-100) containing protease inhibitor cocktail (Sigma, St. Louis, MO, USA). Proteins isolated from HaCaT cells after treatment with PAR-2 AP were separated by SDS-PAGE on 15% gels, and then transferred to nitrocellulose membranes using standard procedures. The membranes were blocked with 5% v/v skim milk in a TBS-T buffer (TBS with 0.1% w/v Tween-20) and reacted with primary antibodies overnight at 4℃. Primary antibodies against PAR-2 (Santa Cruz Biotechnology), cathelicidin (Abcam), and VEGF (Santa Cruz Biotechnology, Santa Cruz, CA, USA) were used at a 1:1000 dilution. The membranes were then washed three times for 15 min each with the TBS-T buffer and incubated for 1 hr with 1% skim milk in TBS-T buffer containing horseradish peroxidase-conjugated secondary antibodies (1:2000). The proteins were visualized using an enhanced chemiluminescence system (Thermo Scientific, Bremen, Germany). GAPDH (Santa Cruz Biotechnology) was used as a control for protein loading.
Statistical analysis
Statistical analyses were performed using Statistical Package for the Social Sciences (SPSS Inc., Chicago, IL, USA) for Windows. Staining intensities of PAR-2 and cathelicidin were compared between normal and rosacea-affected skin using the Student's t-test. The relationship between PAR-2 and cathelicidin staining intensities was tested by Pearson correlation analysis. Differences in PAR-2, cathelicidin and VEGF expression between treatment with PAR-2 AP and PAR-2 CP were analyzed with the Student's t-test. Significance was analyzed at the level of 0.05.
Go to :
RESULTS
Demographic and clinical findings of patients with rosacea
Among the 40 patients with rosacea enrolled in this study, 10 were male and 30 were female. Their ages ranged from 21 to 76 years (mean, 47.2±13.8 years). The physician classified each patient by rosacea subtype and graded the severity of their disease. Twelve patients (30%) were diagnosed with erythematotelangiectatic rosacea (ETR) and 28 patients (70%) were diagnosed with papulopustular rosacea (PPR). No patients presented with the phymatous subtype. Of the 40 patients, six patients (15%) were graded as mild, 24 (60%) were graded as moderate, and 10 (25%) were graded as severe, based on the severity of erythema (Table 1).17
Immunoreactivity for PAR-2 and cathelicidin between rosacea skin and normal facial skin and the correlation between PAR-2 and cathelicidin staining in rosacea tissues
Histopathologically, vascular dilation with perivascular and perifollicular lymphohistiocytic inflammation of varying degrees was noted in the upper dermis of lesional skin from patients with rosacea. Mean staining intensities of PAR-2 and cathelicidin in rosacea were 1.7±0.6 and 1.8±0.7, respectively. Mean staining intensities of PAR-2 and cathelicidin in normal skin were 1.3±1.0 and 0.5±0.3, respectively. Cathelicidin expression in rosacea skin tissues was significantly higher than that in normal tissues (p<0.001). Rosacea tissues showed detectably greater PAR-2 expression than normal skin tissues, although the difference did not reach statistical significance. Additionally, differences in staining intensities between PAR-2 and cathelicidin according to clinical severity were not significant (Table 2). However, a significant correlation between clinical severity and grade of inflammation was observed (p=0.018). As well, differences in expressions of PAR-2 and cathelicidin between subtypes of rosacea, such as ETR and PPR, were also not significant.
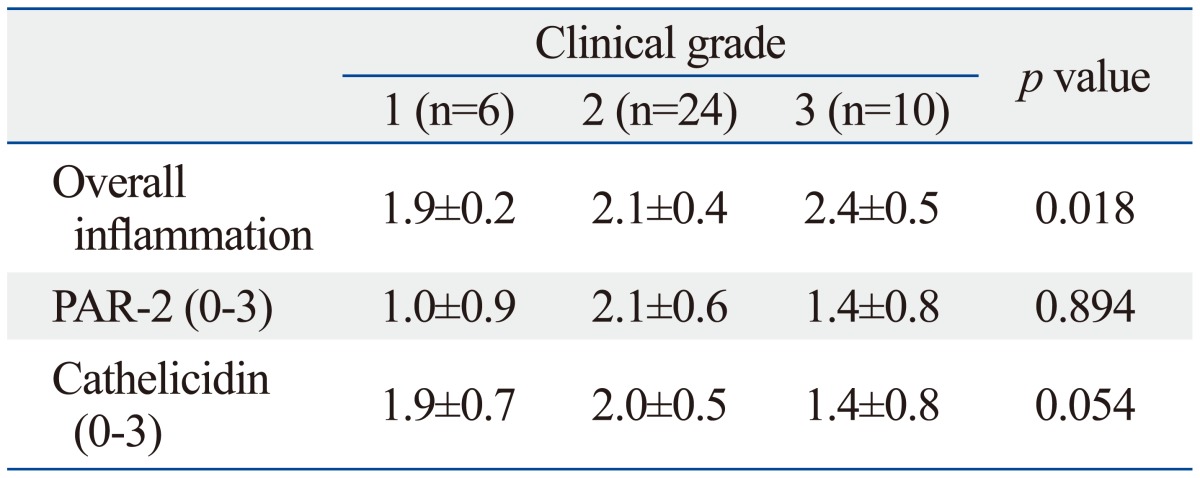
The level of cathelicidin expression according to PAR-2 expression was examined. As PAR-2 expression increased, a trend toward an increase in cathelicidin expression was shown (beta=0.330, p=0.037) (Fig. 1). In Figs. 2 and 3, two patients with rosacea who showed high expression of PAR-2 and cathelicidin in affected skin tissues are presented together with their immunohistochemical findings.
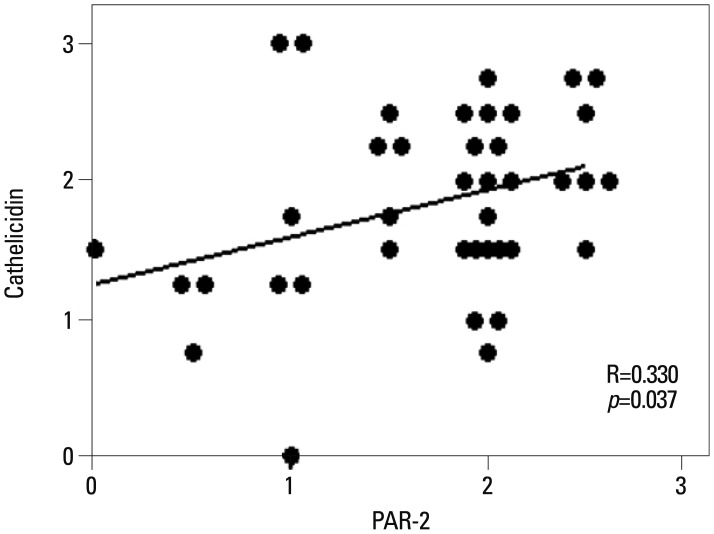 | Fig. 1Significant correlation was observed between PAR-2 and cathelicidin expression in rosacea (R=0.330, p=0.037). PAR-2, protease-activated receptor-2.
|
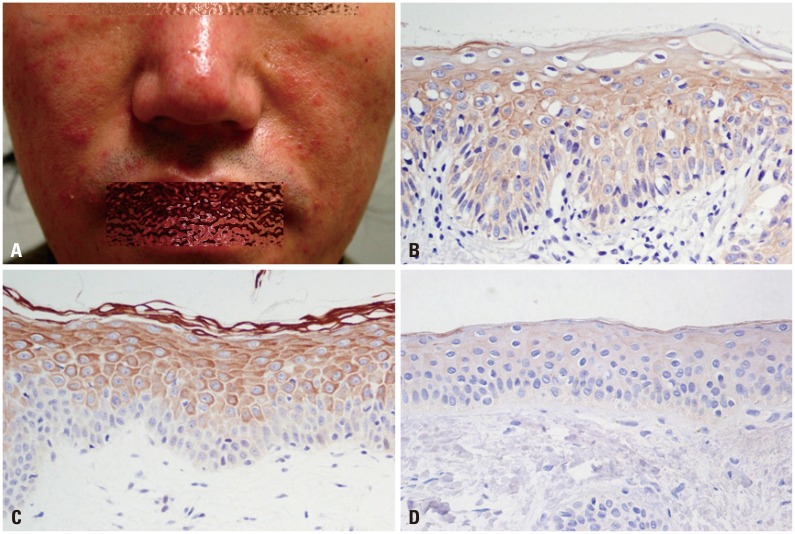 | Fig. 2Clinical picture of a 38-year-old male patient with papulopustular rosacea (A) and immunohistochemical findings of PAR-2 (B) and cathelicidin (C) (H&E, ×400). Staining intensity of PAR-2, 2.5; cathelicidin, 2.5. Cathelicidin staining on normal skin tissue (staining intensity, 0) (D). PAR-2, protease-activated receptor-2.
|
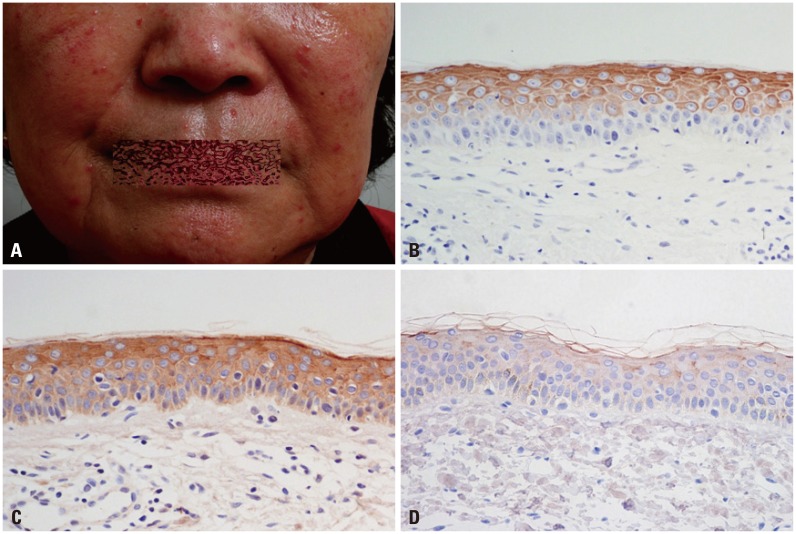 | Fig. 3Clinical picture of a 69-year-old female patient with papulopustular rosacea (A) and immunohistochemical findings of PAR-2 (B) and cathelicidin (C) (H&E, ×400). Staining intensity of PAR-2, 2.5; cathelicidin 3. Cathelicidin staining on normal skin tissue (staining intensity, 0) (D). PAR-2, protease-activated receptor-2.
|
Expression of PAR-2, cathelicidin and VEGF in keratinocytes after treatment of PAR-2 AP
After 24-hr incubation of keratinocytes with PAR-2 AP, the levels of PAR-2, cathelicidin, and VEGF mRNA were significantly higher than those treated with PAR-2 CP (p<0.05) (Fig. 4). In Western blotting, increased levels of PAR-2, cathelicidin, and VEGF proteins were observed after a 24-hr treatment with PAR-2 AP (Fig. 5).
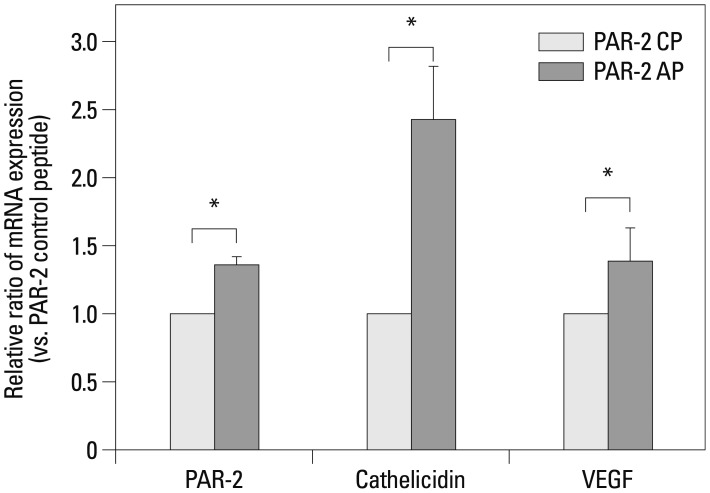 | Fig. 4Effect of PAR-2 AP on the mRNA expression of PAR-2, cathelicidin and VEGF in HaCaT cells. Real time RT-PCR of PAR-2, cathelicidin and VEGF in HaCaT cells after PAR-2 activating peptide and PAR-2 control peptide treatment. Each data point represents the mean (±SEM) result from three independent experiments. *p<0.05. AP, activating peptide; CP, control peptide; VEGF, vascular endothelial growth factor; PAR-2, protease-activated receptor-2; SEM, standard error of the mean.
|
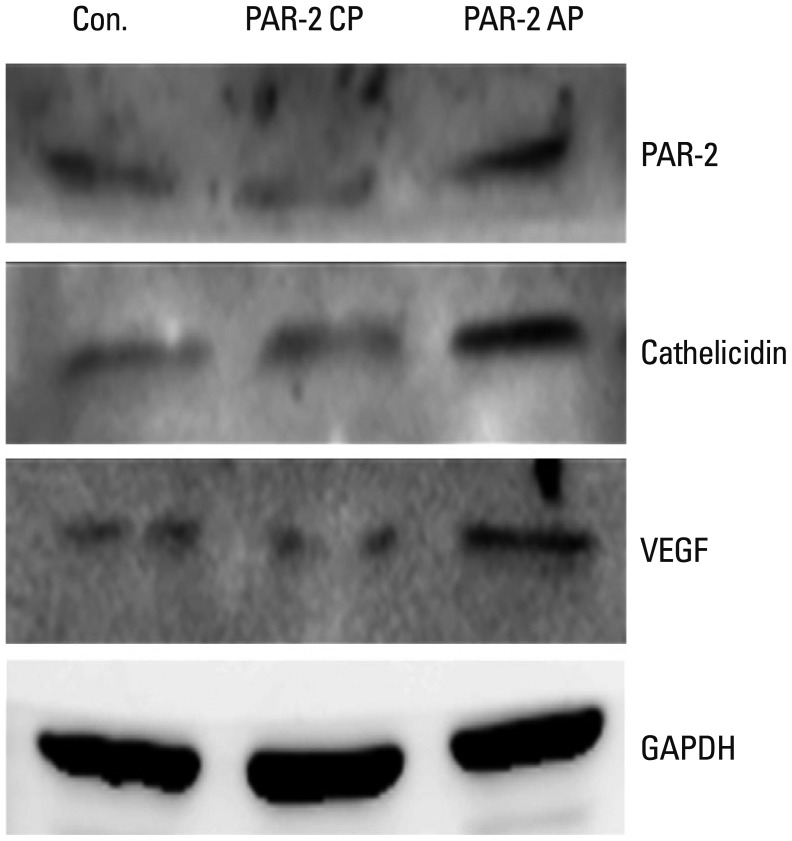 | Fig. 5Effect of PAR-2 AP on the expression of PAR-2, cathelicidin and VEGF proteins in HaCaT cells. Western blotting against PAR-2, cathelicidin and VEGF in HaCaT cells after PAR-2 activating peptide and PAR-2 control peptide treatment. AP, activating peptide; CP, control peptide; GAPDH, glyceraldehyde phosphate dehydrogenase; PAR-2, protease-activated receptor-2; VEGF, vascular endothelial growth factor.
|
Go to :
DISCUSSION
The complex pathophysiology of rosacea suggests a multifactorial etiology with vascular and immunological components underlying a chronic inflammatory response.18 Prominent vasodilation in the affected skin may explain the persistent erythema.19 Microbial agents, notably bacteria associated with Demodex folliculorum infestation, are suspected to activate immune responses in patients predisposed to rosacea.20,21 Altered adaptive and innate immune responsiveness and enhanced cellular reactivity are hypothesized. Abnormal innate immunity is suggested by detection of an AMP and its processing serine protease at higher levels in rosacea lesions than in normal skin.8 In the present study, we tested the involvement of PAR-2 in the expression of AMPs that may contribute to rosacea.
Two important and well-studied types of AMPs in human skin are defensins and cathelicidin LL-37.22,23 hBD-2, the first AMP characterized in human skin, is induced in skin inflammation and infection, and is most effective against gram-negative bacteria.24 Human cathelicidin LL-37, active against both gram-positive and gram-negative bacteria, may be detected directly in human skin only at sites of inflammation.25 Additional functions of LL-37 include chemotactic activation and modulation of cytokines and inflammatory mediators such as cyclooxygenase-2, pro-interleukin (IL) 1b, and IL-8 in keratinocytes.26 Aberrant LL-37 expression at inflammatory sites modifies dendritic cell differentiation, thereby linking innate and adaptive immune responses. The LL-37-activated dendritic cells may potentiate expression of co-stimulatory molecules and a Th1 response in vitro.27 In cultured endothelial cells, LL-37 induces angiogenesis and triggers the proliferation of vessel-like structures.6 Effects induced in vitro by LL-37 correspond to changes seen in rosacea, and altered cathelicidin expression is observed in rosacea-affected skin.8 Although the regulation of cathelicidin LL-37 production is incompletely understood, proteolytic processing of a precursor protein is essential for its activation as a pro-inflammatory molecule.28 The abundance of LL-37 in the lesions of rosacea may result through abnormal production or activity of an epidermal serine protease or of kallikrein, which process the cathelicidin precursor.8,9 Cathelicidin LL-37 is subject to regulation by various factors, including cytokines, bacterial metabolites such as butyrate, hypoxia, and exposure to the sunlight through up-regulation of human cathelicidin antimicrobial protein (hCAP18) expression by vitamin D3.29,30,31,32
The PARs comprise a subfamily of G protein-coupled, 7-transmembrane domain receptors.12,33 PAR-1, -3, and -4 are activated by thrombin, whereas PAR-2 is activated by trypsin-like serine proteases such as kallikrein. Epidermal cells produce several kallikreins, including kallikrein 5 and kallikrein 14, which activate PAR-2.14,34 Detection of PAR-2 and kallikrein 14 in rosacea lesions has been widely reported.35 From this we hypothesized that direct activation of PAR-2 in rosacea lesions may increase cathelicidin expression.
Initially, we found significantly higher cathelicidin expression in rosacea skin tissues than in normal skin, although PAR-2 expression did not differ significantly between normal skin and rosacea. Cathelicidin expression also showed a significant positive correlation with PAR-2 expression on immunohistochemical staining. These findings may plausibly reflect an interaction between PAR-2 and cathelicidin in the pathogenesis of rosacea. Additionally, both cathelicidin and PAR-2 receptor mRNA and protein increased in keratinocytes treated with PAR-2 AP in vitro. These results suggested that not only PAR-2 expression but also cathelicidin could be regulated by direct activation of PAR-2. That is, PAR-2 may serve to trigger an inflammatory response through induction of cathelicidin in keratinocytes. However, as a difference in staining intensities of PAR-2 and cathelicidin according to inflammation and clinical severity was not observed in our study, in the future, further studies regarding relationship between cathelicidin expression and inflammation induction are needed.
PAR-2 AP induces intercellular adhesion molecule-1 expression in human keratinocytes by activating nuclear factor-kappa B.36 Activation of PAR-2 in keratinocytes may also increase production of IL-6, granulocyte-macrophage colony-stimulating factor and IL-8/CXCL 8, promoting granulocyte and T-cell recruitment.37,38 In addition, a reduction of ear swelling and inflammatory infiltrates in PAR-2 (-/-) mice used in a model of contact hypersensitivity indicates a pro-inflammatory role for PAR-2 in allergic dermatitis.39 Just as PAR-2, which is constitutively expressed in epithelial cells, mediates inflammation in diverse tissues, so may PAR-2 present a potentially effective target for the treatment of inflammatory disease.
Interestingly, keratinocytes treated with PAR-2 AP in vitro increased expression of PAR-2 itself. To our knowledge, PAR-2 AP-induced PAR-2 expression has not been previously reported. While PAR-2 activation may stimulate various signaling cascades, including the activation of inflammatory cytokines, conversely, inflammatory cytokines or growth factors, such as IL-1β, TNF-α, TGF-β, and platelet-derived growth factor may stimulate PAR-2 expression.40,41 We therefore considered the possibility that PAR-2 activation by exogenous PAR-2 AP proceeds through a specific cascade leading to PAR-2 expression.
Contrary to expectation, PAR-2 expression did not differ significantly between normal and rosacea-affected skin. Unlike cathelicidin, PAR-2 is constitutively expressed in normal keratinocytes, and judging from immunohistochemical staining results, does not seem to be affected by increased serine protease activity, which is highly expressed in rosacea patients. However, despite the lack of statistical significance, PAR-2 expression in rosacea-affected skin was higher than that in normal skin.
Our findings are limited in that we could not directly compare cathelicidin and PAR-2 expression between lesions and non-lesions within rosacea patients, as this would involve an invasive procedure with cosmetic risks. Similarly, the activity of serine protease was not evaluated because frozen skin tissues are needed for such assay.
Based on our results, we postulated that increased expression of PAR-2 and serine proteases induced by exogenous irritants and aggravating factors may lead to production of cathelicidin itself through PAR-2 signaling and to excessive LL-37 production through processing by serine proteases and that both pathways contribute to the pathology of rosacea. In conclusion, PAR-2 may contribute to the pathogenesis of rosacea through regulatory action of innate immune response. Molecular antagonists of PAR-2 present a plausible therapeutic intervention for rosacea.
Go to :
 XML Download
XML Download