

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Dendritic cell-based tumor immunotherapy became a promising approach in the past decade by the generation of large numbers of immunopotent dendritic cells (DCs) in vitro.1,2 Furthermore, identification of major histocompatibility complex (MHC) class I-associated tumor antigens that can prime antigen-specific CD8 T cells has the potential to facilitate tumor-targeted DC-based vaccine strategies for systemic cytotoxic T lymphocyte (CTL) responses.3,4 While these studies analyzed the efficacy of antigen cross-presentation by DC to CD8+ T cells via MHC class I molecules, optimal systemic anti-tumor immunity and immunological memory involves priming of both CD4+ and CD8+ T cells recognizing tumor-associated antigens.5,6 MHC class II-restricted activation of antigen-specific CD4+ helper T cells are essential for DC maturation that is functionally competent to prime naïve CD8+ T cells.7,8,9,10 Thus, utilization of DC as successful tumor vaccines requires both MHC class I and II-restricted epitopes.
However, the limited availability of tumor-specific MHC class I and II-restricted peptide antigens is still one of the major obstacles.11,12 At present, the great majority of tumors, including pancreatic cancer, are beyond the realm of peptide-based immunotherapy. Accordingly, researchers explored an alternative approach to DCs loaded with peptides by using DCs loaded tumor lysate as a source of tumor antigens to induce protective antitumor responses.13,14 While DCs loaded with tumor lysates exhibited modest protective immunity in various animal tumor models,15,16 and provided limited objective clinical responses in a number of clinical trials,17,18,19 some concerns have been raised about the relative immunogenicity of different modes of tumor lysate preparation.20,21,22,23 Previously, we and others found that heat-treated tumor lysate (HTL) was a better approach for antigen-delivery and tumor immunity compared to conventional tumor lysate (TL) in inducing CD8+ T cells to differentiate into cytotoxic T cells in vitro as well as in vivo murine pancreatic cancer,24 colon cancer,25 or medullary thyroid carcinoma.26 Heat treatment is known to enhance the immunogenicity of tumor cells, which is in part ascribed to heat shock proteins (HSPs), and HSP-chaperoned proteins and peptides released from tumor cells may target DCs with the assistance of HSP receptors and can be taken up by antigen presenting cells (such as dendritic cells and macrophages) through receptor-mediated endocytosis.27 While this approach shares key aspects of tumor lysate in antigen diversity as well as polyclonal CD4 and CTL responses, heat-treated tumor lysate provides additional advantages in stimulating DCs during antigen delivery and possibly in expanded repertoire of presented antigens. Although enhanced delivery of more diverse antigen by this strategy was speculated, quantitative or qualitative analysis of antigen profiles on MHC molecules upon application of this approach has not been performed.
In the present study, we showed that HTL significantly enhanced maturation of DCs through upregulation of antigen-presenting molecules as well as costimulatory molecules. Furthermore, we found that there are qualitative differences in profiles of antigenic peptides eluted from DCs pulsed with TL or HTL from same cell sources by the power of tandem mass spectrometry. While we detected several over-presented peptides in cancer cells, some peptides were identified only in HTL, indicating that HSPs facilitate transfer of specific sets of antigenic peptides onto MHC molecules of DCs. The approach described herein provides a powerful identification method of naturally processed tumor-associated peptides that can aid to formulate tumor-specific vaccines for clinical use.
MATERIALS AND METHODS
Cells and reagents
Panc-1 (human pancreatic ductal adenocarcinoma cell line) and NCI-N87 (human gastric carcinoma) obtained from American Type Culture Collection (Manassas, VA, USA) were maintained in RPMI 1640 supplemented with 20 mM HEPES, pH 7.2, 1 mM sodium pyruvate, 2 mM glutamine, and 10% heat-inactivated fetal calf serum (Life Technologies, Grand Island, NY, USA). The recombinant human cytokines (GM-CSF, IL-4, IL-1β, IL-6, and TNF-α) were purchased from Peprotech (Rocky Hill, NJ, USA) and PGE2 was from Sigma Chemicals (St. Louis, MO, USA).
Preparation of tumor lysate
TL was prepared according to the protocol described by Schnurr, et al.15 Briefly, Panc-1 cells at 90% confluency were digested with 0.02% Trypsin-EDTA and washed once with PBS. After cell counting, cells resuspended in serum-free medium were disrupted by 4 freeze (liquid nitrogen) and thaw (37℃ water bath) cycles. Large particles were removed by centrifugation (10 min, 500×g), and supernatants were passed through a 0.2 µm syringe filter (Pall Corp, Ann Arbor, MI, USA). The protein content of the lysate was determined and aliquots were stored at -80℃. For the HTL generation, Panc-1 cells at 70% confluency were heat-treated for 2 hr at 42℃. Cells were allowed to recover for 24 hr at 37℃ prior to detachment and lysate preparation. Lysates were tested for bacterial endotoxin contamination with the Limulus amoebocyte lysate assay according to manufacturer's instruction (Charles River Endosafe, Charleston, SC, USA) and found to contain less than 0.01 EU/µg protein.
Western blot analysis
Samples of TL and HTL were separated by 10% SDS-PAGE and transferred onto Immobilin PVDF membrane (Millipore, Bedford, CA, USA). After blocking with blocking reagent (Roche Diagnostics, Manheim, Germany), HSPs were detected using antibodies against HSP70, HSC-70, HSP90, and gp96 (StressGen Biotechnologies, Victoria, Canada), followed by anti-mouse IgG HRP (Santa Cruz Biotechnology, Santa Cruz, CA, USA). Specific bands were developed using ECL (Amersham Biosciences, Buckinghamshire, UK).
Generation of monocyte-derived DC and tumor lysate pulsing
All human subjects participated in this study after providing informed consent that was reviewed and approved by the Internal Review Board of Yonsei University College of Medicine. Peripheral blood mononuclear cells (PBMC) were isolated from heparinized venous blood of healthy volunteers by using Ficoll-Hypaque density centrifugation. Monocytes were isolated using CD14 microbeads (Miltenyi Biotech, Auburn, CA, USA), and DCs were differentiated from monocytes as described.28 Briefly, monocytes were cultured in complete RPMI 1640 medium containing GM-CSF (100 ng/mL) and IL-4 (10 ng/mL). On day 6, cells were incubated with TL or HTL for 4 hr. Lysate pulsing was stopped by changing the medium. Subsequently, DC maturation was induced for additional 48 hr by adding a cytokine cocktail consisting of TNF-α (10 ng/mL), IL-1β (10 ng/mL), IL-6 (1000 U/mL), and 10-7 M PGE2.
Flow cytometry analysis
The phenotype of DCs was examined at the start of the culture and after DC maturation (day 8). Monoclonal antibodies against the following antigens were used: CD1a, CD83, CD86, HLA-DR, CD40, and CD80 (all from BD Biosciences, San Diego, CA, USA). Cells were collected and analyzed using Cytomics™ Flow Cytometer (Beckman Coulter, Fullerton, CA, USA). Non-relevant antibodies of recommended isotypes were included as controls. Data analysis was performed by WinMDI (Scripps Institute, La Jolla, CA, USA) or CXP and FCS 3.0 software for FC500 (Beckman Coulter). Cells were electronically gated according to light-scatter properties to exclude cell debris or dead cells stained with propidium iodide (Sigma). Results were expressed as median fluorescence intensity.
Allogeneic mixed leukocyte reaction assay
DCs were harvested, washed, and resuspended in RPMI 1640+10% FBS. A total of 2×105 allogeneic T cells were incubated with irradiated DCs (30 Gy) at different responder/stimulator ratios ranging between 10:1 and 1280:1 in 96-well flat-bottom plates. After 4 days of coculture, the cells were pulsed with [3H] thymidine (1 µCi/mL final concentration) and incorporation was measured after 16 hr. Responses were reported as mean of triplicate counts per min (cpm)±SEM.
A parallel set of cultures was used to determine IFN-γ expression. After 5 days of cultures at 10:1 responder/stimulator ratio of T cells (1×106 cells/mL), T cells were harvested, washed extensively and expanded with IL-2 (10 U/mL, Peprotech) for 6 days. On day 11, T cells (5×105 cells/mL) were harvested and restimulated with IL-2 (10 U/mL) for 24 hr on 24-well plates coated with anti-CD3 antibody (1 µg/mL, BD Pharmingen, San Diego, CA, USA). Samples were assayed in triplicates. The culture supernatants were collected and frozen at -20℃ until measurement.
ELISA assays
IL-12 (p70) and IFN-γ expression levels were determined by a sandwich ELISA assay (OptEIA, BD Biosciences, San Diego, CA, USA). For IL-12 measurement, the cell-free supernatants from immature DCs (day 6) or DCs pulsed with tumor lysate (TL or HTL) and matured by cytokine cocktail for 48 hr were collected. Samples were assayed in triplicates. The assays were carried out according to the instructions provided by the manufacturer. The analysis of significance was carried out using unpaired Student t-tests or one-way ANOVA.
HLA typing of dendritic cells
Genomic DNA was extracted from cultured dendritic cells using QIAamp DNA mini kit (Qiagen, Valencia, CA, USA). DNA-based HLA typing was performed by Luminex Multi-Analyte Profiling system (xMAP) with a WAKFlow HLA typing kit (Wakunaga, Hiroshima, Japan). Briefly, the highly polymorphic exons 2 and 3 of HLA-A, -B, and -C genes and exon 2 of HLA-DRB1 were amplified by polymerase chain reaction (PCR) with biotinylated primers. The PCR product was hybridized with probes immobilized on fluorescent coded microsphere beads. At the same time, biotinylated PCR product was labeled with streptavidin-conjugated R-phycoerythrin and immediately examined with the Luminex (Luminex, Austin, TX, USA). Genotype determination and data analysis were performed automatically using the WAKFlow Typing software according to the manufacturer's instructions.
MHC-associated peptide elution and purification
Peptides were extracted from the surface of 1×107 cells of TL-DC or HTL-DC pelleted in a 15-mL polypropylene tube (Falcon, Oxnard, CA, USA) as described by Santin, et al.29 with minor modifications. Briefly, 2 mL of citrate-phosphate buffer was added directly to pelleted cells and incubated for 5 min at room temperature. After centrifugation at 500×g for 3 min, supernatant was collected and further clarified by centrifugation at 1800×g for 5 min at 4℃. Peptides were stored at -80℃ until further processing.
Each MHC-associated peptide eluted from TL and HTL was purified using an Oasis HLB cartridge (Waters, Milford, MA, USA), lyophilized using a Modulspin 40 from BioTron (Seoul, Korea), and sequentially resuspended in 100 µL of 2% CH3CN in water for nLC-ESI-MS-MS analysis. Peptides were stored at -80℃ before use.
Nanoflow LC-ESI-MS-MS
The nanoflow LC-ESI-MS-MS system utilized was CapLC equipped with Q-TOF Ultima mass spectrometer (Waters, Milford, MA, USA). For nanoflow reversed phase liquid chromatography (nRPLC or nLC), eluted peptides from TL-DC or HTL-DC was separated using a homemade pulled tip capillary column [75 µm inner diameter (i.d.)×360 µm outer diameter (o.d.)×15 cm long] which was packed with a methanol slurry of Magic C18AQ (3 µm, 100 Å) resins from Michrom BioResources Inc. (Auburn, CA, USA) in our laboratory. Prior to RPLC run, each peptide sample was first loaded to a trapping column (200 µm i.d.×360 µm o.d.×1.0 cm long) for on-line sample desalting. The trapping column was prepared by packing Magic C18AQ (5 µm, 200 Å) resins in capillary tubing ended with frit (2 mm in length) which was made by a sol-gel preparation. The trapping column and analytical column were connected via a PEEK microcross, and Pt wire was used as an electrode to supply electric voltage for the electrospray ionization as described in literature.30
A 10 µL aliquot of each eluent from TL-DC or HTL-DC was injected via autosampler to the trapping column, followed by a binary gradient separation as previously reported.31 Briefly, 2% acetonitrile (buffer A) and 95% cetonitrile in water containing 0.1% formic acid (buffer B) were used for binary gradient nLC runs. The flow rate for nLC run was adjusted to 200 nL/min and binary gradient run conditions were as follows: buffer B increased from 2 to 15% for 15 min, then linearly increased to 12% over 10 min, to 32% over 60 min, ramped to 80% over 3 min, and then it was held at 80% for 10 min to clean the RP column. Finally, buffer B was decreased to 2% and was held for 20 min to re-equilibrate the column. Eluted peptides from the analytical capillary column were directly introduced into MS via electrospray ionization (ESI) in positive ion mode at an ESI voltage of 1.8 kV.
Screening and identification of MHC-peptide complexes
MS analysis of MHC-peptide ions was carried out with a precursor scan (300-1800 atomic mass unit, amu) followed by 3 data-dependent MS-MS scans. Collected MS-MS spectra of peptides obtained from collision-induced dissociation experiment were analyzed using the Mascot Search program against NCBI human databases. The mass tolerance between measured monoisotopic mass and calculated mass was 1.0 amu for precursor ions and 0.5 amu for fragment ions. Peptides were accepted only if a MOWSE score higher than 50 and E-value less than 0.05 were satisfied when screening search data. In addition, no fixed and variable modifications and no enzymatic cleavage were selected, including doubly miss cleavages.
RESULTS
Heat treatment enriched HSPs in HTL
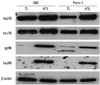
Tumor cell lysate potentially containing multiple tumor-associated antigens are most frequently exploited for DC-based tumor immunotherapy, particularly in cases where suitable tumor-associated antigens are not defined. In our previous study, we showed that DCs pulsed with tumor lysate preparation after heat treatment (HTL) significantly enhanced the antitumor immune responses compared to that of TL. One possible explanation for the superiority of HTL to their counterpart (TL) in transferring antigens onto DC in vitro as well as in antitumor immunity in vivo is the fact that HSPs are involved. HSPs are molecular chaperone of nascent polypeptides including antigenic peptides generated inside the cell. Upon release from dying cells, these peptide-bound HSPs are recognized and endocytosed by antigen presenting cells (such as DCs) via receptor-mediated endocytosis. The molecular interaction has been reported to stimulate antigen presenting cells to secrete proinflammatory cytokines and mediate DC maturation.32 To verify the expression of HSPs resulting from heat-treatment of Panc-1 cells, TL and HTL were subjected to immunoblot analysis. For heat treatment, Panc-1 and N87 cells at 70% confluency were incubated for 1 hr at 42℃. Upon culture for 24 hr at 37℃, cells were evaluated for HSP expression, using antibodies against the most prominently studied HSPs (Fig. 1). Expression of inducible HSP70, gp96, and HSP90 shown to elicit antitumor immunity32 increased dramatically in HTL, while the expression of HSC70 remained relatively unchanged. These data indicate that HTL of Panc-1 contains inducible HSP families that may play a pivotal role in augmenting tumor antigenicity.
Heat-treated tumor lysate (HTL), but not conventional tumor lysate (TL), induced maturation of dendritic cells
To examine whether HTL might affect the expression of surface markers involved in maturation, immature DCs (iDCs) were incubated with TL, HTL (at 100 µg/mL), or media for 4 hr. After pulsing, iDCs were cultured with GM-CSF+IL-4 in the presence or absence of cytokine cocktail (TNF-α, IL-1β, IL-6, and PGE2) for an additional 48 hr to assess the effect of tumor lysate on DC maturation. Finally, nonadherent cells were harvested, stained with antibodies against MHC class II molecules (HLA-DR), CD1a, CD83, CD86, CD40, and CD80 (Fig. 2). For clarity, mean fluorescence intensities of each staining are shown in Fig. 2. The cytokine cocktail induced DC maturation as evident from up-regulation of CD83, HLA-DR, costimulatory molecules (CD80 and CD86), and CD40 as compared to controls. Surface expression of HLA-DR and CD86 was upregulated 2- and 2.1-folds. When cultured in the presence of HTL, even iDC showed signs of maturation, i.e., there was increased expression of HLA-DR (18.6%), CD86 (8.8%), and CD40 (17.7%) on cell surfaces. While TL enhanced the expression of HLA-DR (10.5%), CD40 (7.1%), CD83 (15.6%), and CD80 (12.0%) only marginally, the expression of CD86 (-9.9%) was significantly downregulated by TL. When DCs were stimulated with cytokine cocktail upon lysate loading, the effect of HTL on the expression of maturation markers were more significant. While HTL upregulated the expression of HLA-DR (34.3%), CD40 (9.4%), CD83 (43.0%), CD86 (12.7%), and CD80 (11.9%), TL reduced the expression of HLA-DR (-28.8%), CD86 (14.9%), and CD80 (18/8%). These results imply that TL treatment of DCs decreases T cell activation capacity of either iDC or mature DC (mDC). Yields and viabilities of mDCs were not significantly different from each other (data not shown).
To evaluate the allostimulatory capacities of DCs loaded with TL or HTL, we performed allogeneic mixed leukocyte reaction assay. As shown in Fig. 3A, HTL-loaded iDCs led to stronger allogeneic T cell proliferation compared to that of unpulsed iDCs. Enhanced proliferation of responder T cells was observed in all DC/T cell ratios tested. While allostimulation by TL-loaded iDCs was comparable only to iDCs, DC loaded with HTL and matured with cytokine cocktail (HTL-DCs) were more effective in stimulating a proliferative response than that with TL-DC or control. There was no significant difference in allostimulatory capacity between mDC (control) or TL-mDC. These data show that HTL is more effective in inducing phenotypic and functional maturation of dendritic cells than TL, suggesting that the function of enriched HSPs in HTL may be involved in the induction of DC maturation.
Since IL-12 plays a crucial role in Th1 priming, we then examined the level of IL-12 production by DCs pulsed with TL or HTL. While iDC produced low level of IL-12, maturation with proinflammatory cytokine consistently increased IL-12 production. The production of IL-12 was significantly enhanced in HTL-DC compared to that of unpulsed DC, while no significant difference between TL-DC and HTL-DC was observed (Fig. 3B). We further evaluated the nature of primary allogeneic T cell responses stimulated by mature unpulsed DC, TL-DC and HTL-DC. In line with IL-12 production capacity, HTL-DC significantly stimulated the secretion of IFN-γ from allogeneic T cells compared to that of TL-DC and unpulsed DC (Fig. 3C).
To rule out the possibility that the capacity of tumor lysate to induce DC maturation was due to mycoplasma contamination, as proposed by others,33 lysates were tested for the presence of different mycoplasma strains and proved to be negative (data not shown).
Differences in HLA-associated peptide repertoire between HTL-DC and TL-DC
For the identification of MHC-restricted potential tumor antigens as well as tumor biomarkers on DCs pulsed with tumor antigens, we utilized a MHC-associated peptide proteomics that allows the identification of HLA-restricted peptides on as little as 1×106 dendritic cells. Thus, we exposed dendritic cells to TL or HTL derived from Panc-1 cells and stimulated them with the cytokine cocktail. Dendritic cells were allowed to ingest tumor lysate for 48 hr, whereupon HLA-bound peptides were eluted from dendritic cells with mild acid and analyzed by nLC-ESI-MS-MS.
Furthermore, to explore whether there are any qualitative differences between TL-DC and HTL-DC, the whole HLA-associated peptide repertoire of DCs was investigated. Thus, MHC-associated peptides were sequentially filtered through a 10-kDa cut-off device, and fractionated by RPLC on a capillary C18 column. Fig. 4 shows base peak chromatograms (BPCs) on MHC-associated peptides derived from TL-DC and HTL-DC. The 2 BPCs appear to be significantly different from each other, indicating that HTL-DC displays distinct-peptide repertoire compared to that of TL-DC. HLA genotype of the DCs employed for the peptide elution in the present study was HLA-A*2402, 2901, B*0705, 2705, Cw*0102, 1505, and DRB1*0101, 0803.
Table 1 shows a list of peptides that were present on TL-DC and HTL-DC. Among 200 identified peptides, the list include only those present in 2 independent measurements of TL and HTL-pulsed dendritic cells. Subsequent evaluation and selection by several quality criteria (see Materials and Methods) led to identification of 194 peptides derived from 64 different proteins (Supplementary Table 1, only online). Proteomic profiling of human monocyte-derived dendritic cells revealed that a number of these peptides are derived from ubiquitously expressed proteins (e.g., from house-keeping, cytoskeletal, or matrix proteins).34,35,36,37,38,39,40,41,42,43 These include ribosomal proteins, actin and actin-related proteins, aldolase A, annexin A2, β2-micoglobulin, calmodulin, cathepsin proteins, superoxide dismutase, systatins, elonase 1, galectin 1, glycolysis-related enzymes, histones, HSP70, IL-1RA, kynureninase, macrophage migrating inhibitory factor, peroxiredoxin 6, phosphoglycerated kinase 1, S100 calcium binding proteins and vimentin. Others are derived from proteins that have been shown to be overexpressed in certain cancers, like thymosin beta 4,44 L-plastin,45 makorin-1,46 transmembrane glycoprotein nonmetastatic melanoma protein B (gpnmb),47 cofilin48 or S100A4.49 As a representative example of shared peptides between TL-DC and HTL-DC, the candidate ion of m/z 828.32 (M+2H+)2+ of TL-DC was found to have peptide sequence DKDGNGYISAAELRHVM belonging to human S100 calcium binding protein A4 (residues 94-110, NCBI accession number gi|75765831). The identical peptide was also identified from m/z 625.94 ion ([M+3H]3+) from HTL-DC-eluted samples (Fig. 5).
Identification of pancreatic cancer-related antigenic peptides presented on the surface of HTL- and TL-DC
In the list, there are a number of peptides that are known to be associated with pancreatic cancer including galectin 1,50 cofilin,48 oncogene DJ-1,51 coactosin-like 1,52 cathepsin,53 and S100A454 (Table 1). Exactly the same peptides could also be detected in an independent experiment in which the same dendritic cells were pulsed with different batches of TL and HTL from Panc-1 cells (data not shown), suggesting that epitopes are HLA-restricted and very abundant on MHC class I or II molecules of dendritic cells after uptake of TL or HTL derived from Panc-1 cells. While most of the peptides listed were present in both eluents from HTL-DC and TL-DC that are derived from either ubiquitously expressed proteins (i.e., housekeeping gene products) or from endogenously processed proteins from dendritic cells, MARKS, makorin-1 (zfn-xp protein), CECR-1, thyroid adenoma-associated protein isoform b and transmembrane glycoprotein nonmetastatic melanoma protein B (gpnmb) were detected only in eluents from HTL-DC. On the other hand, prothymosin alpha and cofilin 1 were detected only in TL-DC. Furthermore, comparison of peptides shared by HTL-DC and TL-DC revealed that there are delicate differences in the list of peptides from same proteins. For example, shorter peptides from AHNAK nucleoprotein isoform 1 (desmoyokin) and gelsolin were more abundant in HTL-DC than TL-DC, suggesting that these peptides are derived from class I HLA molecules via cross-presentation. While association with other types of tumor has been reported, AHNAK, SH3 domain binding glutamic acid-rich protein like 3, calpastatin, hnRNP-E1, thyroid adenoma-associated protein isoform b and gpnmb were new additions to pancreatic cancer-associated proteins. Among these, makorin-1, CECR1 and thyroid adenoma-associated protein isoform b protein were previously unrecognized for pancreatic and/or other types of tumors.
DISCUSSION
One of the challenges in DC-mediated cancer immunotherapy is how to load tumor-associated antigens (TAAs) to DCs in order to generate effector T cells that are powerful enough to overcome systemic and local immunosuppression imposed by tumor. Strategies for antigen-loading to DC include synthetic antigenic peptides,55 freeze/thaw lysate,13 apoptotic tumor cells,56 DC-tumor cell fusion,57 and DC priming with tumor RNA.58 The anti-tumor activity of tumor cell lysate-pulsed DCs was first reported in 1998 and was utilized to treat malignant diseases including melanoma, prostate cancer, renal cancer, lung cancer, lymphoma, brain tumor, liver cancer, and colorectal cancer.59 Theoretically, whole tumor priming strategies providing full repertoire of tumor-specific and tumor-associated antigens are advantageous, as they lower the chance of tumor evasion of the immune response by antigen deletion or mutation and circumvent the need for identification of TAAs from individual cancers. In an effort to further enhance the anti-tumor effect of whole tumor cell lysate-pulsed DCs, we reported previously that tumor lysate prepared from heat-treated tumor cells containing abundant chaperoned TAAs generated a more potent immune response by eliciting higher IFN-γ secretion, and stronger CTL response with greater tumor growth retardation against non-immunogenic murine pancreatic ductal adenocarcinoma as compared with tumor lysate.24 While this approach shares key aspects of tumor lysate in inducing polyclonal cellular immune responses, heat-treated tumor lysate provides additional advantages in stimulating DCs during antigen delivery and possibly in expanded spectrum of presented antigens, resulting in a broader repertoire of elicited T-cell responses.
What mechanisms could account for the higher efficiency of HTL in antigen delivery and DC activation over TL? We favor the concept that composition of HTL greatly differs from TL in the variety and abundance of molecules they contain, thereby making available for antigen presenting cell (APC). It was shown that heat stress induces the expression of HSPs that can carry peptides and proteins derived from tumor-associated proteins60 and are efficiently taken up by APCs with the assistance of the cell surface HSP receptors,61,62,63 helping the antigenic peptide presentation on MHC-I as well as promoting the function of antigen-presenting cells, especially dendritic cells, to generate antigen-specific CD8+ T cells.64,65,66 Thus, HSP-chaperoned TAAs in HTL may represent specific fingerprints of tumor cells which are different from conventional TL. Furthermore, heat-induced HSPs are known to serve as endogenous danger signals to drive DC maturation.67,68 Our findings that the immunostimulatory capacity of heat-treated tumor lysate-loaded DCs (HTL-DC) is more potent than from tumor lysate-loaded DC (TL-DC) clearly supports the postulation.24 Compared to TL-pulsed DCs, immunization with HTL-pulsed DCs induced a large amount of specific CTLs with higher tumor specific cytotoxic activity in naïve mice in vivo. Accordingly, various strategies utilizing various formats of HSPs are currently being tested as vaccines in clinical trials against cancer.65,69 Herein, we addressed the question of whether these 2 types of lysate preparation differ in their capacities to promote maturation of DC and in their antigenic peptide repertoire available for DCs to present to T cells.
HTL, but not TL, induced partial DC maturation in the absence of maturation factors and this effect was further enhanced by the maturation factor (cytokine cocktail), as shown by DC phenotype, allostimulatory capacity and IFN-γ secretion from allogeneic T cells. Thus, increased HSPs in tumor cells may serve as chaperons for antigenic peptides and natural adjuvants for DC maturation during the process of DC antigen processing, presentation and enhancing naïve T cell stimulatory capacity of DCs. The ineffectiveness of TL in stimulating DC suggests that DCs are not responsive to constitutive HSP70 family proteins (HSC70). As HSP is known to facilitate the transfer of antigenic proteins or peptides from tumor lysate to DC, it is likely that peptides chaperoned by HSPs in TL may be taken up by DC with lower efficiency than that of HSPs in HTL. As reported recently, upregulation of HSP expression by cell stress elicited more potent anti-tumor immunity.70,71
In an effort to delineate differences in antigenic peptide profiles from HLA class I and II molecules on DCs pulsed with TL or HTL, we analyzed HLA-associated antigenic peptides derived from dendritic cells through a highly sensitive and well established tandem mass spectrometry (MS-MS) method. Even with this approach, however, only a fraction of the peptides in a complex sample can be successfully sequenced due to the limited dynamic range of MS instrumentation. To identify low abundant tumor cell-derived peptides among the total mixture of MHC class I and II peptides presented by DCs, including self-peptides and tumor-derived housekeeping proteins, peptides identified were compared to previously reported proteomic database of human monocyte-derived dendritic cells. From a single Panc-1 tumor line, more than 194 peptide ligands present in these 2 sets of DCs were identified. The peptides identified varied in size, ranging 8-29 amino acids. In general, MHC class I presents short peptide sequence (8-12 amino acids) and MHC class II present much longer peptides of 14-25 amino acids. However, there are longer peptides that can bind and be eluted off the MHC class I molecules by a protrusion mechanism.72,73 For example, the repertoire of peptides bound to a subset of HLA-B27 in vivo revealed the molecular mass range of 900 to 4000 Da (8 to 33 amino acid residues), unlike the major HLA-B27 with 900-1400 Da (8-12 amino acids). In agreement with the previous report of Weinschenk, et al.,74 the majority of peptides presented in MHC class I and II on the surface of DCs were from common housekeeping proteins (proteins associated with metabolic processes, cell growth and protein synthesis). Apparently, MHC-associated Panc-1-derived ligands contained numerous self-peptides reflecting a comprehensive summary of proteome of normal cells.75
Other peptides were derived from proteins that are overexpressed in pancreatic cancer. Examples in this category were L-plastin,45 profilin 1,76 cofilin 1,48,76 DJ-1,77 AHNAK,78 calpastatin,79 and oncogene prostate tumor inducing gene-1.80 We also identified proteins that have not previously been correlated with pancreatic cancer, including gpnmb, makorin-1 (znf-xp protein) and CECR1 protein (cat eye syndrome critical region protein 1 precursor). As DCs function as vehicles for antigen presentation to the immune system, characterization of MHC peptide antigens from tumor lysate-pulsed DCs would greatly facilitate the identification process of immunogenic peptides for tumor immunotherapy. These peptides in turn could serve as disease markers and indicators of tumor progression.
A most intriguing observation of this study was that the identified peptides eluted from DC upon co-culture with HTL and TL were not identical. First of all, there were differences in profile identified proteins from eluted peptides (i.e., repertoire). While some proteins (makorin-1, CECR1, thyroid adenoma-associated protein isoform b and gpnmb) were detected only in HTL-DC, other proteins (prothymosin alpha and cofilin 1) appeared exclusively in TL-DC. Furthermore, even in proteins shared by HTL-DC and TL-DC, peptides eluted from HTL-DC contained smaller peptides (8-12 amino acids) that were not present in eluents from TL-DC. This finding strongly suggests that DCs cross-presented HTL via receptor-mediated endocytosis (such as by HSPs) more efficiently than conventional TL.
In the present study, we performed the first successful comparative analysis of HLA-associated peptides, which are derived from TL and HTL-pulsed dendritic cells, by nLC-ESI-MS-MS, resulting in a sensitive and rapid sequencing of naturally processed HLA-associated peptides. To the best of our knowledge, a comparative study on HLA-presented tumor antigens, TL-DC and HTL-DC, by using LC-MALDI-MS and LC-ESI-MS has not previously been published. We believe that the approach introduced here provides a powerful complementation to the current methods for expanding the number of human tumor antigens. Vaccine strategies aim to inducing both CD4 and CD8 T cell responses with appropriate combinations of identified tumor antigens may contribute to increase the success rate of cancer immunotherapy.





 XML Download
XML Download