

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Although the incidence and mortality of gastric cancer have decreased in the Western world, gastric cancer persists as a common malignancy and leading cause of cancer-related death in Asian contries.1,2 Surgery is the only curative treatment for gastric cancer. Chemotherapy achieves favorable results for unresectable advanced and recurrent gastric cancers; however, the prognosis for these cancers is very poor.
Taxanes such as paclitaxel and docetaxel are a class of anticancer agents that bind to the β tubulin subunit of polymerized microtubules and induce hyperstabilization, which causes cell cycle arrest and apoptosis.3 Taxanes are used most commonly for the treatment of breast, lung, ovary, and gastric cancer. The taxanes paclitaxel and docetaxel exhibit similar efficacies in gastric cancer treatment.4-11
There are at least eight mammalian β tubulin isotypes, and the isotype composition in humans varies across tissues.12 Overexpression of class III β tubulin (TUBB3) has been observed in several human cancer cell lines such as prostate, ovarian, breast, and non small cell lung cancer. Cells expressing TUBB3 show resistance to docetaxel and paclitaxel.12-15 A small cohort study of advanced gastric cancer patients who were receiving preoperative docetaxel-based chemotherapy detected a correlation between expression of TUBB3 and poor response to chemotherapy.16
Hypoxia in solid tumors is associated with resistance to chemotherapy, induction of angiogenesis, and poor patient prognosis, and angiogenesis is a hallmark of human malignancies. The induction of vascular endothelial growth factor (VEGF), mediated by interacting genetic and environmental signals, is an essential component of tumor angiogenesis.17
The transcription factor hypoxia-inducible factor-1α (HIF-1α) is a primary regulator of VEGF during hypoxic conditions, and Calvani, et al.18 reported that inhibition of VEGF or the VEGF receptor-2 (VEGFR-2), abolishes VEGF-mediated induction of HIF-1α. That is, HIF-1α increases VEGF induction, and VEGF also induces HIF-1α expression. A recent report suggested that HIF-1α also mediates TUBB3 induction in hypoxia.19-21 Through these results, we postulated that blockade of VEGF in gastric cancer cells would be associated with a decrease in HIF-1α and TUBB3 expression and a concomitant increase in sensitivity to paclitaxel.
The response of gastric cancer cells to anti-VEGF antibodies is of interest because bevacizumab, a humanized monoclonal antibody against VEGF, is used currently as an anti-angiogenic treatment for metastatic colorectal cancer, non-squamous non-small cell lung cancer, and metastatic breast cancer.22-24 Recent the Avastin in Gastric Cancer trial evaluated the efficacy of adding bevacizumab to chemotherapy in the first-line treatment of advanced gastric cancer. Adding bevacizumab to chemotherapy was associated with significant increases in progression free survival and overall response rate, but not associated with significant increases in overall survival.25 To date, we had insufficient evidences for the benefit of adding bevacizumab to chemotherapy in the treatment of gastric cancer.
Placental growth factor (PlGF) was discovered shortly after VEGF. Whereas VEGF binds both VEGFR-1 and VEGFR-2, PlGF binds to VEGFR-1 but not VEGFR-2. The key role of VEGF and its receptor VEGFR-2 in tumor angiogenesis is firmly established, but the contribution of VEGFR-1 remains poorly defined.26 In the present study, we investigated the relationships between VEGF, HIF-1α, and TUBB3 in gastric cancer cells (AGS). We report that blockade of VEGFR-1 and VEGFR-2 with concomitant paclitaxel treatment increase the cell cytotoxicity of TUBB3-expressing gastric cancer cells.
MATERIALS AND METHODS
Drugs and reagents
Paclitaxel (BMS Pharmaceuticals, New York, NY, USA) and bevacizumab (Roche Pharmaceuticals, Basel, Switzerland) were diluted in absolute DMSO. This solution was diluted to 0.1% DMSO on each day of experimentation. The PlGF and VEGFR-1/2 neutralizing monoclonal antibodies were purchased from research and development systems.
Cell culture and hypoxic treatment
The human gastric cancer cell lines, AGS, SNU, and TMK1 were used as in vitro model systems. Cells were cultured in RPMI 1640 with 10% fetal bovine serum. For normoxia, cultures were incubated at 37℃ in a humidified chamber of 5% CO2. For hypoxia, cells were treated with CoCl2. Confirmatory experiments under hypoxic conditions (1% O2/5% CO2/94% N2) were performed using a Modular Incubator Chamber-101. Each hypoxia experiment was conducted under CoCl2-treated and true hypoxic conditions, and the results were equivalent.
siRNA
The TUBB3 siRNA duplex targeted a sequence 531 bases downstream of the TUBB3 start codon (siTUBB3: 5'-CACGGTGGTGGAGCCCTACAA-3'). siRNA duplex oligonucleotides were purchased from QIAGEN (Gaithersburg, MD, USA) and transfected at 100 nM using Lipofectamine RNAiMAX Reagent (Invitrogen, Carlsbad, CA, USA).
Cell proliferation assay
To estimate viability, 105 cells were seeded in 24-well plates, subjected to normoxia or hypoxia, and exposed to neutralizing antibodies in the culture media for 4 h. Cells then were treated with bevacizuamb and/or paclitaxel for 24 h. Cells were incubated for 24, 48, or 72 h, and EZ-CyTox (Daeil Lab Service, Seoul, Korea) was added to each well. After 3 h, absorbances at 420 nm were measured using a microplate reader (EL800, BioTek, Winooski, VT, USA).
Tubulin polymerization assay
Cells were treated with various concentrations of drugs for 24 h, washed twice with phosphate buffered saline, and lysed for 5 min in 100 µL hypotonic buffer [1 mM MgCl2, 2 mM EGTA, 0.5% NP40, 20 mM Tris-HCL (pH 6.8), protease and phosphatase inhibitors]. Samples then were centrifuged at 16000×g for 10 min at room temperature. Supernatants, containing soluble tubulin, were separated from pellets, containing assembled tubulin. Each fraction was separated by SDS-PAGE. Immunoblots then were performed using anti-pan-α-tubulin rabbit polyclonal antibody (Santa Cruz Biotechonology, Santa Cruz, CA, USA).
Immunoblotting
Cells were lysed in Mammalian Cell Lysis Reagent (Fermentas, Waltham, MA, USA), and approximately 30 µg protein was separated in 8-10% polyacrylamide gels and transferred to polyvinylidene difluoride membrane (Millipore, Billerica, MA, USA). The following antibodies were used: anti-HIF-1α and anti-TUBB3 (Abcam PLC, Cambridge, UK), anti-tubulin (Invitrogen, Carlsbad, CA, USA), β-actin (Santa Cruz Biotechnology, Santa Cruz, CA, USA).
RESULTS
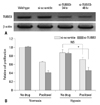
Induction of HIF-1α, TUBB3 and VEGF in hypoxia
The expression levels of TUBB3 and VEGF were measured during the induction of HIF-1α in hypoxic AGS. AGS exhibited well-defined hypoxic induction of HIF-1α, VEGF, and TUBB3 compared with other cell lines (SNU638, TMK1) (Fig. 1).
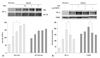
TUBB3 confers gastric cancer cells with resistance to paclitaxel
To assess whether TUBB3 expressions is associated with chemotherapeutic drug resistance in AGS, TUBB3 was knocked down with small interfering RNA (siTUBB3), and the effects of paclitaxel on AGS growth under normoxic and hypoxic conditions were evaluated over a 24-48 h period. AGS was strongly resistant to paclitaxel during hypoxia. Knockdown of TUBB3 decreased the resistance to paclitaxel during hypoxia, and cytotoxicity was increased (Fig. 2).
VEGFR-1 signals increase HIF-1α and TUBB3 expressions in AGS cells
After treatment with paclitaxel, cytoskeletal component were isolated from AGS cells, and tubulin polymerization was monitored. Western blots were performed using a pan α-tubulin polyclonal antibody to probe lysates from cells that had been treated with paclitaxel or paclitaxel and bevacizumab under normoxic conditions or after 24 h of hypoxia. Paclitaxel activity was inhibited in hypoxia. Tubulin polymerization was decreased in hypoxia compared with normoxia and it was statistically significant (p<0.01). Hypoxic decrease of polymerization was lessened after treated with bevacizumab (Fig. 3A).
As previously mentioned, we postulated that blockade of VEGF in AGS would be associated with a decrease in HIF-1α and TUBB3 expression and a concomitant increase in sensitivity to paclitaxel. We treated cells with bevacizumab, anti-VEGFR-1 and anti-VEGFR-2 neutralizing antibodies to test this hypothesis. AGS cells were cultured under normoxic and hypoxic conditions for 24 h without exogenous growth factors, in the absence or presence of anti-VEGFR-1, anti-VEGFR-2, or bevacizumab with simultaneous paclitaxel treatment. HIF-1α and TUBB3 were increased in a hypoxic condition. Paclitaxel treatment decreased the hypoxia-dependent increased expression of HIF-1α and TUBB3. When paclitaxel was combined with bevacizumab or anti-VEGFR-2, expression levels of HIF-1α and TUBB3 were further decreased. The greatest decrease in HIF-1α and TUBB3 levels occurred in response to paclitaxel and anti-VEGFR-1 treatment in AGS cells (Fig. 3B). Our results using AGS cells contrast with results using HCT116 cells, in which hypoxia-induced HIF-1α was inhibited via VEGFR-2.18
To examine VEGFR-1 function, we investigated whether the VEGFR-1 ligand, PlGF, influences the expression of HIF-1α in AGS and HCT116 cells. Cells were cultured under normoxic or hypoxic conditions in the presence of PlGF (range, 0.1-2.0 ng/mL). HIF-1α expression was increased in hypoxia and was dose dependently more increased according to the concentration of PlGF (Fig. 4A). In contrast with AGS cells, PlGF rarely influenced HIF-1α expression in HCT116 cells. AGS cells were cultured under normoxic and hypoxic conditions in the absence or presence of PlGF and anti-VEGFR-1. Expression levels of HIF-1α and TUBB3 were increased in hypoxia and were more increased in response to PlGF treatment. The increased expressions of HIF-1α and TUBB3 were suppressed when anti-VEGFR-1 antibodies were combined with PlGF treatment (Fig. 4B). We concluded that induction of HIF-1α in AGS is VEGFR-1 mediated and PlGF dependent.
Blockade of VEGFR-1/VEGFR-2 increases sensitivity to paclitaxel in AGS cells
AGS cells were cultured under hypoxic conditions with or without anti-VEGFR-1, anti-VEGFR-2, and bevacizumab with simultaneous paclitaxel exposure. The cell viability assay demonstrated that AGS cell cytotoxicity was increased in response to combined treatment with paclitaxel and bevacizumab compared with paclitaxel treatment alone. Cytotoxicity was increased synergistically in response to simultaneous treatment with paclitaxel, anti-VEGFR-1, and anti-VEGFR-2 (p<0.05) (Fig. 5).
DISCUSSION
Bevacizumab is accepted as the standard treatment for advanced metastatic colorectal cancer, but to date is not considered as the standard treatment of gastric cancer.24 VEGF can bind with the receptor tyrosine kinases, VEGFR-1 and VEGFR-2. The key roles of VEGF and its receptor VEGFR-2 in tumor angiogenesis have been established.26,27 Activation of VEGFR-2 leads to auto-phosphorylation and the activation of downstream signaling pathways, such as Raf/MEK/ERK and PI3K/Akt kinase cascades.25 In contrast, the function of VEGFR-1 remains poorly defined. VEGFR-1 has been associated with tumor growth, tumor cell activation, and metastasis. Treatment with VEGFR-1 inhibitors, such as anti-VEGFR-1 antibody, suppresses tumor growth and metastasis in various models.26 However, the degree of tumor growth inhibition varies. Some studies have reported that VEGFR-1 inhibition is not sufficient to block tumor growth without combined inhibition of VEGFR-2.28,29 In addition, it is unclear if VEGFR-1 inhibitors, by preventing the binding of VEGF to VEGFR-1, increase the concentration of free VEGF that can subsequently activate VEGFR-2 and stimulate angiogenesis by an alternate pathway. The precise mechanisms of VEGFR-1 inhibitor functioning warrant further exploration.26
We demonstrated that HIF-1α expressions increase in association with elevated PlGF. HIF-1α and TUBB3 were upregulated in hypoxia and became more induced in response to PlGF treatment. We also demonstrated that inhibition of VEGFR-1 is associated with decreases in HIF-1α and TUBB3 expression. Calvani, et al.18 reported that VEGFR-2, rather than VEGFR-1, mediates VEGF-dependent induction of HIF-1α in hypoxic HCT116 colon cancer cells, suggesting that functional VEGFR-2, but not functional VEGFR-1, exists in this colon cancer cell line. Cell viability assay results demonstrated that AGS cell cytotoxicity was most pronounced when cells were treated simultaneously with paclitaxel, anti-VEGFR-1, and anti-VEGFR-2. Through these results, we suggest that AGS cells express functional VEGFR-1 and VEGFR-2.
TUBB3 confers chemoresistance to taxanes. Our experiment results of paclitaxel resistance in AGS cells are consistent with other reports suggesting that TUBB3 affects chemoresistance to taxanes in diverse cancer cells.12-16 Raspaglio, et al.21 also reported that TUBB3 is not only a parameter related to the chemoresponsiveness, but also could be a pure prognostic marker. TUBB3 expression levels differ by cell and tissue types. In some tissues, TUBB3 is constitutively expressed and is not inducible upon hypoxia.21 In AGS cells, we observed TUBB3 was constitutively expressed in normoxia and became more induced in hypoxia.
The present study demonstrated that blockade of both VEGFR-1 and VEGFR-2 in conjunction with paclitaxel synergistically depresses induction of HIF-1α and TUBB3 expression, and more effectively increases cytotoxicity in gastric cancer cells.



 XML Download
XML Download