

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Influenza viruses continue to change their antigenicity by successfully evading the host immunity acquired by previous vaccinations or natural exposures to infections and claim the lives of 250,000 to 500,000 people worldwide annually. The antigenic drift by genetic mutations in the influenza viral genome not only leads to the emergence of antiviral drug-resistant strains,1,2 but also evades antibody-mediated viral neutralization (VN).3 Vaccination remains the most cost-effective countermeasure against influenza virus infection. The effect of vaccination relies on the induction of high levels of neutralizing antibodies specific to the viral surface proteins, hemagglutinin (HA) and neuraminidase (NA), which mediates the initial attachment of the virus to specific cellular receptors on the cell surface promoting the entry of the virus into the target cell and the cleavage of the sialic acid moiety from the receptor permitting the release of progeny virus particles, respectively. Influenza vaccines are prepared each year with the aim of matching the strains that would circulate in that season. The current inactivated influenza vaccines contain three distinctive antigens from two influenza A virus strains (H1N1 and H3N2) and one B strain as recommended by the World Health Organization (WHO). While providing protective immune responses against homologous and closely related virus strains, they rarely protect against antibody-escape variants of seasonal influenza viruses or newly circulating strains in subsequent influenza seasons, hence necessitating an annual update of new HA and NA antigens. Furthermore, as seen in the previous outbreaks of the highly pathogenic avian H5N1 influenza virus (HPAI), a completely new strain against which contemporary vaccines provide little or no protection could emerge from non-human avian reservoirs.4-6 Although a sustained human-to-human transmission of the H5N1 HPAI has not yet been observed, it should be noted that three major influenza pandemics of the 20th century were all caused by influenza A viruses originating from birds, calling for constant vigilance and monitoring. Recent demonstration that guided mutations in the HA of the HPAI markedly increased the transmission efficiency in mammalian hosts partially mirrors similar antigenic drifts in nature as well.7-9 Meanwhile, the sudden emergence and rapid transmission of the 2009 pandemic H1N1 influenza virus (pdmH1N1) was recorded as the first pandemic in the 21st century. The pdmH1N1 itself was shown to have a unique genetic constellation that has not been previously reported.10,11 This raised a possibility that further genetic reassortment of the virus with other virulent strains (antigenic shift) could generate other novel pandemic strains with a high virulence and human transmissibility. In support of this hypothesis, experimental evidence showed a considerable genetic compatibility between the HPAI and the pdmH1N1, where the reassortants between the two viruses led to high transmission ability among mammalian hosts.12,13
The persistent threats posed by antigenically diverse and rapidly evolving influenza viruses heighten the interests in the development of cross-protective influenza vaccines and ultimately a universal vaccine that could provide protection against antigenic drift and shift strains of influenza. Relevant to this issue, the cross-reactive cytotoxic T lymphocytes (CTLs) and broadly neutralizing antibodies directed to the conserved domains of influenza viral proteins have been proposed as the most likely triggers to eliciting cross-protective immunity across different influenza strains. T cell-mediated cross-reactivity is frequently observed in influenza infections among different strains of the same virus and, in rare occasions, even among unrelated viruses such as hepatitis C virus and Epstein-Barr virus.14 Numerous reports on influenza viruses have demonstrated that the T cell responses are crucial for viral clearance by removing the virus-infected cells from the host.15-18 However, the CTL responses are considered a 'double-edged sword' since, while providing cross-protective immune responses, they sometimes aggravate disease symptoms by having a pathogenic effect.19-21 However, broadly neutralizing antibodies have recently emerged as reliable effectors for the cross-reactivity against the influenza viruses; also providing a promising avenue to the development of cross-protective influenza vaccines. The two major principles underlying cross-reactivity provide the cornerstone for developing cross-protective influenza vaccines.
In light of cross-protection, cold-adapted live attenuated influenza vaccines (CAIVs) are of much interests since they can mount all phases of immune responses, including systemic and local humoral responses as well as cell-mediated immunity. Along with the CTLs and the broadly reactive antibodies, the intranasal administration of the CAIVs stimulates the induction of secretory IgA antibodies in the respiratory tract, which crucially contributes to cross-protection.
Here, we review the experimental and clinical evidence for cross-reactive immunity afforded by the CAIVs, and further discuss possible mechanisms underlying the cross-protection by the vaccines. In addition, important considerations for the development of more effective and cross-protective influenza vaccines are described; therefore, tipping the balance of the CTL responses from a pathogenic to a beneficial cross-protective immune response.
INACTIVATED INFLUENZA VACCINES AND CROSS-PROTECTION
Inactivated influenza vaccines have been most widely used in preventing influenza infection in humans for more than fifty years. There are two types of inactivated vaccines currently used clinically. Split-virus vaccines are prepared by disrupting the whole virions containing all viral particles and the ssRNA genome, whereas subunit vaccines contain the highly purified HA and NA proteins-the two most immunogenic surface glycoproteins. Inactivated influenza vaccines preferentially induce serum IgG antibodies directed to HA, which neutralize the virus by binding the globular head domain of the HA and preventing the initial attachment of the virus to cellular receptors on the surface membrane.
In general, inactivated vaccines are poorly immunogenic, requiring at least two separate vaccinations and appropriate adjuvants to induce sufficiently protective immune responses.22,23 Since protection by the vaccines highly depends on the induction of the neutralizing antibodies against the HA, the vaccines need to be reformulated yearly according to the antigenic changes in a subsequent season. Occasionally, the vaccine strains fail to properly match the circulating strains; therefore, the vaccine effectiveness is significantly reduced, which was documented most apparently during 1997-1998.24,25 Seasonal influenza vaccines could be prepared in advance upon recommendation from the WHO or CDC based on global surveillance. Pandemic outbreaks, however, often accompany an antigenic shift resulting from genetic reassortment between more than two different strains, as seen in the pdmH1N1,10,11 rendering the previous vaccinations completely ineffective against the new pandemic strains.
The diversity of reservoirs, frequent animal-to-human transmission, and highly variable nature of the influenza virus led us to develop alternative vaccination strategies to enhance the broadly reactive antibodies and the CTL responses. For instance, immunization with a novel immunogen comprising the conserved stalk domain of the HA and lacking the variable head domain provided a broad spectrum of protection against diverse virus strains in mice.26 In parallel, alternative vaccine strategies were tested for their potential of improving the CTL responses. Since the CTL responses are directed to antigenic peptides processed in the cytoplasm of the antigen presenting cells, endogenous protein synthesis is essential to inducing functional CTL responses against the influenza viral infection. Immunization with a DNA construct encoding influenza viral proteins leads to in situ synthesis and processing of the viral antigens in the cytoplasm of the injected cells, which are then loaded into the major histocompatiblity complex (MHC) class-I molecules to subsequently stimulate the CTL responses. DNA immunization against influenza antigens, including HA, NP, M1, and M2, has shown protective immunity against homologous and heterologous infections in various animal models,27-29 but safety concerns remain with regard to the use of DNA for mass vaccination purposes.30
With respect to cross-protective immune responses, annual use of inactivated vaccine may hamper the development of the influenza virus-specific cross-reactive CTLs.31-34 In mice and ferrets, the use of inactivated A/H3N2 vaccines prevented the induction of heterosubtypic immunity to a lethal infection with influenza A/Indonesia/5/2005 (H5N1), which was found to correlate with reduced CTL responses.32-34 Similar patterns were also observed in humans, in which children who had received annual influenza inactivated vaccines did not show the age-dependent increase in the frequency of influenza-specific CTLs.31 It was assumed in those reports that the annual vaccinations early in life, particularly with inactivated vaccines, might lead to skewed immune responses preferentially enhancing the humoral responses. Therefore, in those reports, the use of the live attenuated influenza vaccine - able of inducing the virus-specific CTLs as well as the humoral responses - was recommended to the young children, in light of inducing cross-protective immunity to render them protected against antigenic variants or pandemic threats as well.
ADVANTAGES OF THE CAIVS FOR CROSS-PROTECTION
As an attractive alternative to the inactivated vaccines, the CAIVs have been used in humans against annual seasonal influenza virus infections since 2003. In addition, the CAIVs against a pandemic or a potential pandemic strain, such as the pdmH1N1 and H5N1 HPAI, were developed and evaluated for their safety and efficacy in animal models.35-40 Besides inherent issues of safety and efficacy, the CAIV provides specific advantages pertaining to cross-protection. The CAIVs are delivered via intranasal administration mimicking a natural infection by the influenza virus and hence is able to mount the local antiviral immunity, which has been thought to provide cross-protection. Such cross-protective immunity could also be effectively elicited by intranasal delivery even with the inactivated vaccines.41,42 Furthermore, in addition to humoral antibody responses directed to surface antigen proteins, the CAIVs replicate to a limited extent in the respiratory tracts and deliver internal viral components, which are then subjected to antigen presentation pathways involving MHC class-I molecules, consequently stimulating the CTL responses. Taken together, the CAIVs appear to encompass almost all immune responses that would not be expected from inactivated or DNA vaccines. However, the CAIVs may be less competent than inactivated or DNA vaccine in eliciting one particular phase of an immune response. For example, subunit vaccines are composed of highly purified surface antigens; therefore, are able of inducing higher serum IgG antibody titers than the CAIVs. It was reported that the CAIVs induced higher levels of local IgA antibody but lower levels of serum hemagglutinin inhibition (HI) antibody than inactivated vaccines, which became more prominent in the elderly people aged 50 and more, suggesting the use of both vaccines in combination for achieving optimal protection efficacy in the group.43,44 The DNA vaccine encoding internal viral proteins might be more specialized for stimulating the CTLs than the CAIVs, due to the over expression of the selected proteins under the control of a strong eukaryotic promoter such as the cytomegalovirus promoter. However, each of these strategies cannot stimulate the same wide range of immune responses expected from the CAIVs. Among several advantages presented by the CAIVs, stimulation of broad immunity against heterologous strains is of particular interest, especially because it may open exciting opportunity of developing a 'universal vaccine' that confers a wide range of protection against diverse antigenic strains, which remains an ultimate goal in the vaccine research field.
REPORTED EVIDENCE FOR CROSS-PROTECTION BY THE CAIVS
It has been well established that natural infection with an influenza virus results in protective immunity against reinfection with the same virus strain (strain-specific immunity), drift viruses within the same subtype (subtype-specific immunity) and even with different subtype viruses (heterosubtypic immunity), albeit with less strength than against a homologous strain.45 Such commonly observed cross-protection by natural infection serves as a working model of protection afforded by the CAIVs. The CAIVs have been extensively investigated for their genetic stability, immunogenicity, and protective efficacy in a number of preclinical and clinical trials, before becoming licensed for human use as a trivalent formulation containing two A types (H1N1 and H3N2) and one B type strain. To date, a number of reports have shown that immunization with the CAIVs elicits varying degrees of cross-protective immunity to heterologous influenza virus strains (Table 1). Those reports were initially focused mainly on the subtype-specific immunity that was demonstrated within the same subtype, between the vaccine and its drifted variants. In field trials for humans, vaccinations with the CAIVs induced serum HI antibodies against a drifted strain that was not contained in the vaccine, and contributed to a significant reduction of illness by infection with the drift virus, as compared to the placebo recipients.46-49 Subsequently, it was reported that in humans the degree of cross-protection by the CAIVs could vary depending on the age of recipients, with higher efficacy reported in children than adults.50,51 Realizing that the cross-protective immunity depends on the CTLs specific to the conserved epitopes of influenza viral proteins, efforts to illuminate the breadth of the cross-reactivity afforded by the CAIVs were extended to heterosubtypic immunity operating among different subtypes. In mice, cold-adapted H3N2 vaccine provided partial resistance to H1N1 infection through the CTL responses, the depletion of the CTLs cancelling out the protection against the challenge, highlighting the importance of the CTL responses to eliciting cross-protection.52
With the HPAI emerging as serious pandemic threats to human health, several types of contemporary seasonal vaccines were assessed for their cross-reactivity against the HPAI,53-55 and yet there has been no published data addressing the cross-reactivity between the seasonal CAIVs and the HPAI. Heterosubtypic cross-protection against the HPAI conferred by cold-adapted 2009 pdmH1N1 vaccines40,56 indirectly suggests the possible cross-reactivity between them. It is not surprising that immunization with the A/Ann Arbor/6/60 ca (H3N2), the CAIV donor strain itself, provided partial protection against the HPAI infection in mice.35 This result implies again that immunization with the cold-adapted donor strain can provide a certain degree of partial protection against heterologous infections, regardless of the subtype of challenging virus. This basal level of cross-reactivity afforded by the CAIV donor strains should present a practical means to develop the universal vaccine. Relevant to the issue of cross-reactive immune responses, enormous antigenic diversity in the HPAI makes it imperative that a H5N1 vaccine should cover a broad range of H5N1 variants from different clades or subclades.57,58 This cross-clade protection was examined in various animal models including mice, ferrets, and nonhuman primates, in which one or two doses of immunizations with the H5N1 CAIV elicited protective antibody responses against both homologous and heterologous strains with different clade or subclade HPAIs.35,36,59
The sudden emergence and global circulation of the pdmH1N1 with an unprecedented speed was against common prediction that H5N1 HPAI would be the next pandemic strain. Fortunately, pdmH1N1 caused fewer deaths than previous pandemics before moving into its postpandemic period. In parallel with global efforts to develop CAIVs against the pdmH1N1,39,40,60,61 many researchers also evaluated the cross-reactivity between the seasonal CAIVs and the pdmH1N1, with the hope that the seasonal CAIVs that contained H1N1 subtype would elicit protection against the pdmH1N1. The comparative studies with seasonal CAIVs and inactivated vaccines revealed the superior protection against pdmH1N1 by the seasonal CAIVs, and the cross-reactive CTL responses were proposed to be the most likely correlates for this protection.38,62,63 Of note, in humans with prior exposure to the pdmH1N1 or immunized with inactivated H1N1 vaccine, broadly cross-reactive antibodies dominated the human B cell responses against heterologous influenza strains,64,65 suggesting that the HA of the pdmH1N1 might carry many conserved epitopes and was able to preferentially elicit antibodies specific to those epitopes. Such broad reactivity of the HA of the pdmH1N1 was further confirmed in an animal model study, in which immunization with the pdmH1N1 CAIVs conferred high level of cross-protection against the seasonal and the HPAI infections in mice.40,56
MECHANISMS UNDERLYING THE CROSS-PROTECTION BY THE CAIVS
In this section, we discuss currently proposed mechanisms for the cross-reactivity of the CAIVs (Table 2) suggested in the references discussed above, which could be further extended to the cross-reactivity seen in other live attenuated vaccine strategies;66 however, with a varying degree of contribution of each factor depending on the attenuation tools employed.
CTL responses as a general basis for cross-protection
The CTLs are MHC class-I-restricted CD8 T cells that play a central role in killing the host cells infected with viruses or intracellular microbial pathogens.67 Since the CTLs recognize foreign peptides that are degraded in the cytoplasm and loaded into the MHC class I molecules, influenza internal components constitute the primary resources of antigenic peptides pool for the influenza specific CTLs. However, the two surface glycoproteins (HA and NA) are expressed by the ribosomes on the rough endoreticulum reticular and remain embedded in the lipid membrane until translocated to the cytoplasmic membrane. These surface antigens are processed and loaded into the MHC class-II molecules in endocytic vesicle, and then presented to CD4 T cells for stimulation of antibody production by the B cell.67 It has been well recognized that non-glycoproteins are highly conserved according to the type (A, B, or C), and therefore the CTLs specific to a certain influenza strain will elicit a broad spectrum of responsiveness to diverse viral subtypes within the same type. Indeed, several reports showed that the influenza virus-specific CTLs were directed against the NP and M1 protein,18,68,69 based on which the influenza viruses are divided into three types (A, B, and C). Moreover, many studies suggested that the cross-reactive immunity between different subtypes were mediated by the CTLs, as deduced from diverse combinations of priming and challenge experiments in animal models.45 It is now established that the cross-reactive CTLs play crucial roles for viral clearance and the decrease the morbidity associated with infection, although it alone cannot prevent infection. Several studies aimed to identify potently immunogenic and conserved T cell epitopes embedded in influenza viral proteins in addition to NP and M1 and aimed to design more broadly protective vaccines.70-72 As demonstrated in the CAIVs, the CTL responses were demonstrated to be critical factors responsible for eliciting cross-protection against heterosubtypic infections. The pdmH1N1 CAIV protected the immunized mice from heterologous infections with the seasonal H1N1 and the avian H5N1 virus, even without measurable HI or viral neutralization (VN) antibody titer, inducing significant specific CTL responses to the viruses, which suggested the important role of the CTL responses for protection.56 Although the reports summarized in Table 1 does not fully represent the data on the CTL responses, it is highly predictive that the CTL responses would participate in the cross-protection, which would contribute to more significantly to heterosubtypic protection than to subtype-specific or cross-clade protections.
IgA antibody-mediated cross-protection
Administrated via the nasal route, a typical entry site used by the influenza virus, the CAIVs are able to induce local antiviral immunity in the upper and lower respiratory tracts (RT). Of particular importance with respect to the cross-protection is the induction of secretory IgA antibodies (sIgA) in the RT. The sIgA is a major humoral mediator of mucosal immunity and, like IgM, has a tendency to form a polymeric structure that is essential for transport through the epithelium. Moreover, the immunoglobulin polymerization is thought to be associated with higher affinity or binding strength for antigens than monomeric IgG with the same specificities.73,74 This consideration was also relevant to influenza virus-specific sIgA in the RT, because the polymeric sIgA was more cross-reactive with a several-fold higher activity in hemagglutination inhibition and virus neutralization than monomeric IgG.75 In line with these observations, the cross-reactivity of the sIgA was stronger than serum IgG, in mice immunized with the pdmH1N1 CAIV. It was demonstrated that while the cross-reactive serum IgG titers were only a tenth of the homologous strain-specific IgG titers, the sIgA titers in the RT were similar across homologous and heterologous viruses including seasonal and H5 influenza strains.40
Non-neutralizing antibodies
Although neutralizing antibodies are recognized as primary effectors responsible for successful protection against homologous infection, there is also a wealth of evidence that non-neutralizing antibodies are intimately associated with protection, especially against heterologous infections. While the NP-specific CTL responses were recognized to be one of the responsible factors for heterosubtypic resistance, the involvement of NP-specific antibodies in protection has largely been disregarded. This was not because of the absence of the NP-specific antibody but because of its relatively weak contribution to overall protection because the NP-specific antibodies provided only partial protection when passively transferred into naïve mice.76 In another study, the non-neutralizing antibodies by themselves did not provide any protection to heterologous challenge in animal model, however they reduced the morbidity and promoted recovery from the infection in cooperation with memory T cells.77 Intriguingly, the non-neutralizing antibodies were also found to facilitate the expansion of responding memory CD8 T cells, which suggests the coordination of B cells and T cells in eliciting the cross-protective immunity.77 Other protective mechanisms by the non-neutralizing antibodies involve either macrophages or natural killer (NK) cells. IgG antibodies bound to a virus particle are recognized by Fc receptors expressed on the macrophages, which then actively ingest the opsonized virus, playing a pivotal role in the clearance of the infection.78 However, the NK cells were reported to recognize antibodies bound to the influenza M2 proteins expressed on the surface of virus-infected cells and this finally lead to cell death also known as the antibody-dependent cell-mediated cytotoxicity.79 These mechanisms may explain the existence of the non-neutralizing antibodies found in the serum of CAIV-immunized animals that yielded a considerable level of IgG antibody titers in ELISA, while not being detected in HI assay nor in VN assay.40
Antibodies specific to highly conserved domains of HA
From the late 2000s, the highly variable surface glycoprotein, HA, has become the focus in identifying conserved regions in the protein and their corresponding antibodies expected to be cross-reactive, with the aim of developing universal vaccine constructs. These successful findings not only provide a promising avenue for novel vaccine strategy, but also illuminate an additional contributing factor to the cross-protection conferred by the CAIVs. Two conserved regions in the HA were responsible for recognition by newly discovered antibodies - the membrane-proximal stem domain harboring the membrane fusion peptide80-82 and the receptor binding pocket in the globular head domain.83,84 Monoclonal antibodies that recognized each of these two domains provided heterologous immunity when passively transferred into naive mice. These findings also open the possibility of significantly broadening the cross-reactivity of the influenza live vaccines through rational design. This may be possible, for example, via appropriate genetic engineering of the vaccine such that the exposure of the conserved domains of the HA could be enhanced while maintaining other desirable characters. Recently, the structural and genetic basis for such broadly neutralizing antibodies was identified as largely originating from a specific antibody gene, heavy-chain variable region IGHV1-69.65,85 These reports could invigorate further discoveries of hitherto unknown antibodies derived from the antibody gene.
Innate immunity
All of the principles of cross-protection explained above are results from adaptive immune responses acquired mainly after viral infection or vaccination. While adaptive immunity is highly specific to a particular pathogen or its products, innate immunity provides immediate protection against a wide range of pathogens, although the protection is partial and short-lived.67 Innate immunity involves the induction of antiviral interferon responses and pro-inflammatory cytokines and chemokines, which help healthy cells resist pathogens including viruses.67 It has been reported that cold-adapted X-31 (A/H3N2) donor strain provided 100% protection in mice when the vaccine was inoculated three or four days prior to a lethal challenge with either heterosubtypic (A/New Caledonia/99, H1N1) or even a heterotypic (B/Shangdong/97) strain.86 The immediate and broad spectrum of protection was shown to be mediated by innate immunity rather than by specific antibodies. The innate immunity by the CAIVs seems to be beneficial particularly in the event of an unexpected and sudden emergence of an influenza pandemic of a completely new subtype when there is no sufficient time for the production of a matching vaccine. Therefore, the CAIVs provide an immediate and broad-spectrum protection against various influenza strains, which may extend and complement the current influenza control strategies.
IMMUNODOMINANCE IN THE CTL RESPONSES AND PATHOGENIC EPITOPES
As discussed above, a potential drawback associated with the annual use of inactivated vaccines in children comprises a preferential and skewed induction of the humoral responses over the CTLs, raising the concern of decreased cross-protective immunity.87-89 Conversely, an imbalance within the influenza virus-specific CTLs was also recognized as a potentially pathogenic factor leading to the unexpected loss of homologous protective immunity by an exaggerated expansion of the cross-reactive but less protective CTLs. Direct evidence for the existence of such phenomenon has not yet been presented for the influenza virus. Considering that the CAIVs induce strong CTL responses, it is worth discussing closely related cases relevant to the rational design of a more broadly-protective live vaccine without any pathogenic effects.
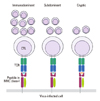
Of the many virus-derived peptides processed and presented, only a few stimulate strong CD8 T cell responses, which means that a hierarchy exists of dominance in the T cell responses, with some peptides recognized strongly (immunodominant), some weakly (subdominant), and others only in the absence of the dominant peptides (cryptic) (Fig. 1).90 It was previously proposed that the hierarchy of immunodominance shaped by previous exposure to a pathogen varies markedly upon a secondary exposure to a heterologous strain. T cells specific to cross-reactive epitopes shared by the two pathogens dominate both the primary and memory pool while those specific for non-cross-reactive epitopes are selectively lost.21 This alteration in the immunodominance hierarchy was described for influenza viruses as well, which lead to a selective expansion of cross-reactive T cells upon subsequent infection with a heterosubtypic influenza strain.91 This raises a reasonable question as to why most people become susceptible to influenza variants circulating in a subsequent season despite repeated prior exposures to the viruses. The most plausible answer to this question may be that, without the help of neutralizing antibody responses, the cross-reactive T cell responses by themselves do not provide sufficient protection against the heterologous influenza strains. This consideration provides strong support towards vaccination with the CAIV because it would boost the underlying cross-reactive T cell responses and induce protective neutralizing antibody responses as well.
Despite the potential advantages of selectively expanding cross-reactive T cell responses by vaccination, serious concerns were also raised by other publications on potentially pathogenic effects of the cross-reactive T cell epitopes. For example, herpes simplex virus-1 and theiler's virus were found to stimulate the autoreactive T cells that target the proteins of the eye and brain, leading to conjunctivitis and encephalitis, respectively.92,93 Although there is no reported evidence yet for the existence of self-reactive T cell epitopes in influenza viral peptides, caution should be taken when selecting and modulating the cross-reactive epitopes for generating more cross-protective live vaccines.
Furthermore, it was shown that if the cross-reactive but less protective CTL response was selectively expanded by subsequent heterologous infections, then the pre-existing normally protective T cell response would become markedly suppressed leading to the attrition of protective immunity to the previous homologous strain.94 Considering that the CTL responses alone fail to support the protection against the influenza viruses, such cross-reactive pathogenic epitopes may raise concern, especially for individuals with pre-existing strong cross-reactive T cells by repeated exposures to the viruses but with poor neutralizing antibodies. In those individuals, vaccination may aggravate, rather than alleviate, the symptoms from the infection. This consideration emphasizes the importance of neutralizing activity for successful protection against the influenza viruses and further suggests that a rational design of influenza live vaccines should focus primarily on its ability to induce neutralizing antibodies against the HA and NA required to prevent infection in the first place, and secondarily on increasing the cross-protection.
CONCLUSION
A hugely diverse antigenicity and the continual evolution of influenza viruses through antigenic drift and antigenic shift represent the biggest challenge to the development of a long lasting and broadly protective vaccine. The CAIVs appear better fit to the concept of cross-protection than inactivated vaccines for several reasons discussed above. In addition to making better use of previously known mechanisms, further discovery and appropriate modulation of novel cross-protective T cell epitopes would greatly improve the protection coverage of the vaccines. Likewise, it will be worthwhile to generate diverse strains of cold-adapted donor strains because each different backbone strain would differentially influence the immune responses both in quantity and quality, particularly through the CTL responses directed to internal viral proteins of the donor strain.
Although some of the principles underlying the cross-protection were extrapolated from previous reports dealing with inactivated vaccines or experimental infections, it is reasonable to assume that factors uncovered from those studies may also contribute to the cross-protection afforded by the live vaccines. In addition to the cold-adaptation, several attempts were made to develop alternative live vaccine strategies by reverse genetics technology.67 With varying degrees depending on their attenuation mechanisms, they will also stimulate a similar repertoire of immunological correlates responsible for cross-protection. However, this does not imply that live vaccines always provide the best options for inducing the highest level of cross-protection. More specialized regimens such as DNA vaccines or other viral vectored vaccines encoding multiple conserved epitopes under a strong promoter should also be considered.
Many studies on cross-protection using infection models or vaccinations revealed a huge complexity of the immune system, ranging from the coordination of multiple cellular and humoral components required for cross-protection, to the T cell immunodominance that in some cases may exert some harmful effects through autoimmunity or the loss of protective immunity by exaggerated expansion of cross-reactive but less protective CTL responses. These findings underscore a more detailed understanding of our immune system and the importance of predicting the evolution of the influenza viruses for the rational design of the universal influenza vaccines.

 XML Download
XML Download