

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Thyroid cancer is the most prevalent endocrine malignancy, and its incidence has been increasing.1,2 Well-differentiated thyroid carcinoma has a favorable prognosis with surgical and radioactive iodine therapy. Other thyroid malignancies such as poorly differentiated thyroid carcinomas and anaplastic thyroid carcinomas frequently metastasize, precluding patients from a curative resection. Therapeutic options for these patients include additional surgery for resectable lesions, external radiotherapy and chemotherapy. The result of this approach is usually disappointing and the use of novel therapeutic approaches is needed. Poorly-differentiated thyroid carcinoma and anaplastic thyroid carcinoma, which are thought to be dedifferentiated forms of pre-existing differentiated tumors, do not respond well to medical and surgical treatment. There are several therapeutic methods, including surgery, radioactive iodine therapy, and thyroid stimulating hormone suppression by thyroid hormone replacement, for treating thyroid cancer. Most thyroid cancers are well differentiated types and can be cured with ease through surgery and radioactive iodine therapy. However, poorly differentiated or anaplastic thyroid cancer shows rapid progression and is difficult to cure.
Berberine, isolated from the roots of herbal plants, is a natural compound that is non-toxic to humans.3 Berberine has been used as an agent for the treatment of inflammatory disease such as rheumatoid arthritis due to its anti-inflammatory activities.4 Berberine also can control several physical activities, including lowering-cholesterol5 and maintenance of insulin levels.6 Anti-cancer activity of berberine against a variety of cancer cell lines has been introduced. Berberine exhibits antitumor effects by inhibiting the growth,7 invasion,8 and metastatic ability9 of several cancers, including Ehrlich ascites carcinoma,10 lung cancer,11 epidermoid carcinoma,12 leukemia,13 cervical cancer14 and breast cancer.15 Though anticancer activity of berberine has been reported in many articles, its effect on thyroid cancer has not been demonstrated.
To observe the effects of berberine on thyroid cancer, we selected two cell lines, 8505C and TPC1. The 8505C cell line has been used to model anaplastic thyroid cancer, and the TPC1 cell line to model papillary thyroid cancer.16
MATERIALS AND METHODS
Cell culture
Unless otherwise specified, all reagents were purchased from Sigma (St. Louis, MO, USA). 8505C and TPC1 cells were cultured on tissue culture dishes (Falcon; Cat no: 353003, San Jose, CA, USA) in Dulbecco's Modified Eagle's Medium (DMEM) containing 10% fetal bovine serum (FBS). For culture, 5×105 cells were cultured at 37℃ in a humidified atmosphere containing 5% CO2.
Assessment of cell growth following berberine treatment
For each cell line, 5×105 cells were seeded on tissue culture dishes in DMEM containing 10% FBS. After 24 hours, the cells were washed with PBS and cultured in fresh media. Berberine (Sigma, Cat no: B3412, 95% purity, Stock solution; 100 mM in dH2O) was added at 1 µM, 10 µM, or 100 µM for 72 hours. Cells were then counted in a Neubauer chamber after incubation with 0.4% trypan blue dye.
Cell cycle analysis
After 10 µM berberine treatment for 72 hours, the cells were harvested, fixed in 70% ethanol for 1 hour, and washed with PBS. Cells were treated with 100 µg/mL RNase A for 1 hour at 37℃, and stained with 10 µg/mL propidium iodide (PI). Flow cytometry was performed in triplicate for each experiment using a FACS Caliber system (BD Biosciences, San Jose, CA, USA).
Apoptosis analysis with annexin V staining
To analyze apoptosis, the cells treated with 10 µM berberine for 72 hours, and then both floating and adherent cells were harvested. Annexin V staining was performed according to the manufacturer's protocol (BD Pharmingen, Franklin Lakes, NJ, USA). Trypsin-EDTA was used to obtain single-cell suspensions. After centrifugation, cells were washed twice with cold PBS and resuspended in binding buffer (10 mM HEPES, pH 7.4, 150 mM NaCl, 5 mM KCl, 1 mM MgCl2, and 1.8 mM CaCl2) at a concentration of 1×106 cells/mL. An aliquot (100 µL) of the solution containing 1×105 cells was transferred to a 5-mL culture tube, and 5 µL each of Annexin V-FITC and PI were added. After vortexing, cells were incubated for 15 minutes at room temperature (25℃) in the dark and 400 µL of 1X binding buffer was added to each tube. Flow cytometry was performed using a FACS Caliber system within 1 hour.
Expression of p27 by immunoblotting following berberine treatment
Cells were detached using trypsin, collected by centrifugation, and cultured as described above. Cells were treated with 10 µM berberine for 72h, and total cell lysates were collected by 200 µL lysis buffer according to the manufacturer's protocol (Cell Signaling Technology, Beverly, MA, USA). To analyze expression of p27, protein concentration was measured using a Bio-Rad Protein Assay kit (Bio-Rad Laboratories, Hercules, CA, USA) according to the manufacturer's directions. Equal amounts of cell lysates were separated by 10% SDS-PAGE. The blocking of blots was processed with a 10% blocking buffer [10% nonfat dry milk in Tris-buffered saline (TBS) containing 0.1% Tween 20] for 1 h. Primary anti-p27 mouse monoclonal immunoglobulin G (IgG) antibody (1 : 1000; Cell Signaling Technology, Beverly, MA, USA) and anti-β-actin mouse monoclonal IgG antibody (1 : 1000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) were incubated overnight at 4℃, and were washed three times in TBS containing 0.1% Tween 20 and secondary peroxidase-conjugated Affinipure rabbit anti-mouse IgG (1 : 2500, Jackson ImmunoResearch Laboratories, West Grove, PA, USA) or peroxidase-conjugated Affinipure mouse anti-rabbit IgG for 1 h at room temperature. Blots of p27proteins were detected with an enhanced chemiluminescence detection system (Amersham Bioscience, Piscataway, NJ, USA).
Statistical analysis
All data were compiled from a minimum of three replicate experiments. Data for statistical analysis were expressed as the mean±standard error. Comparisons of results from treated versus control cells were performed using Student's t-tests. A p-value of less than 0.05 was considered statistically significant.
RESULTS
Berberine treatment inhibited 8505C and TPC1 cell growth in a dose-dependent manner
We previously reported the effects of berberine in a metastatic breast cancer model.15 To observe the effects of berberine on thyroid cancer, we selected two cell lines, the anaplastic thyroid cancer cell line 8505C and the papillary thyroid cancer cell line TPC1. Both cell lines were treated with berberine at several concentrations (Fig. 1). After 72 hours of treatment, growth was inhibited in a dose-dependent manner in both cell lines (Fig. 1). These results show IC50 of berberine at 10 µM in both cell lines. Thus, 10 µM of berberine was selected for the analysis of the cell cycle and Anexin V staining in both cell lines.
Analysis of cell cycle and apoptosis in berberine-treated cells
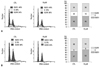
Berberine is known to be an inducing drug for cell cycle arrest and apoptosis. To determine whether growth inhibition was a result of cell cycle arrest, apoptosis, or a combination of both, we analyzed the DNA content of the treated cells by propidium iodide staining and flow cytometry. Growth was inhibited in both cell lines following berberine treatment. In cell cycle analysis using propidium iodide staining, berberine induced a 2% increase in G2/M arrest at 10 µM in 8505C cells (Fig. 2A), whereas TPC1 cells showed a 6% increase in the G0/G1 phase at 10 µM of berberine (Fig. 2B). The cell lines showed different apoptotic responses to berberine treatment. In the Annexin V staining, berberine at 10 µM induced a 7% increase in Annexin V-FITC positive and PI negative 8505C cells (Fig. 3A), but no increase was detected in Annexin V-FITC positive and PI negative TPC1 cells (Fig. 3B).
Expression of p-27 protein in berberine-treated cells
We found that growth inhibition was caused by cell cycle arrest in TPC1 cells and by apoptosis in 8505c cells following berberine treatment. To confirm these results, we analyzed the expression of p-27, which is known as the molecule involved in cell cycle arrest. As shown in Fig. 4, berberine induced a little up-regulation of p-27 in 8505c cells (Fig. 4A) but relatively high up-regulation of p-27 in TPC1 cells (Fig. 4B).
DISCUSSION
Berberine is a natural compound reported to have broad anticancer effects, as well as anti-inflammatory effects. We examined the anticancer effect of berberine on the thyroid cancer cell lines 8505C and TPC1, through examination of cell cycle and apoptosis modulation. We observed that berberine treatment of these cell lines decreased growth rates in a dose-dependent manner. Berberine induced apoptosis in both cell types. However, cell cycle arrest was induced mainly in TPC1 cells.
Our results showed 6% increase of the G0/G1 cell cycle in TPC1 cells and 2% increase of the G2/M cycle in 8505C cells after berberine treatment. These results are consistent with our immunoblotting data in which berberine induced a little up-regulation of p-27 in 8505c cells but relatively high up-regulation of p-27 in TPC1 cells after berberine treatment.
Consistent with our results, it has been reported that cell growth can be inhibited by berberine through cell-cycle arrest.17 Berberine has been shown to inhibit the activity of DNA topoisomerase II alpha17 and Cdki-Cdk-cyclin,12 and to induce G0/G1 cell-cycle arrest in L1210 cells3 and G2/M-phase arrest in leukemia cells.18 Previously, we reported similar berberine-induced inhibition of growth through cell-cycle arrest and apoptosis-induction in breast cancer.15 Together with these reports, our results suggest that the effects of berberine treatment of thyroid cancer cells rely upon the induction of cell-cycle arrest in TPC1 cells. It was already demonstrated in our experiments that berberine inhibited cell growth in a dose-dependent manner in TPC1 cells and induced cell cycle arrest at the G0/G1 phase. It was confirmed by immunoblotting that berberine increased p27 in TPC1 cells. All these results suggest that berberine induced cell cycle arrest by induction of p27, which arrests cells at the G0/G1 phase of the cell cycle in TPC1 cells, and thus that berberine can be a novel candidate for an anti-cancer agent against thyroid cancer. Our results showed that berberine more significantly increased the levels of p27 and was more likely to lead to cell cycle arrest in TPC1 cells with RET/PTC1 rearrangement than in 8505C cells with BRAF V600E mutation. P27 has been shown to be targeted by the BRAF/MEK/ERK signaling pathway in distinct tumors models such as melanoma and thyroid carcinomas.19-21
In addition, previous studies have indicated that berberine could induce apoptosis in several cancers, including epidermoid carcinoma,12 human leukemia HL-60 cells,13 and Ehrlich ascites carcinoma cells.22
Here, we also observed that berberine more strongly induced apoptosis in anaplastic thyroid cancer cell line 8505C than in TPC1. In our experiment, berberine induced an 8% increase of apoptosis in Annexin V-FITC positive and PI negative 8505C cells but, 0% in Annexin V-FITC positive and PI negative TPC1 cells. Since externalization of phosphatidylserine occurs in the earlier stages of apoptosis, FITC Annexin V staining can identify apoptosis at an earlier stage than assays based on nuclear changes such as DNA fragmentation.23 Of course, we cannot confirm apoptosis using FITC Annexin V staining, and DNA Fragmentation assay should be done to clearly identify the cytotoxic or cytostatic activities of berberine. However, these results show that growth inhibition by berberine is not associated with cytotoxic effects but with cell cycle arrest through induction of p27 in TPC1 cells.
In conclusion, we have demonstrated that berberine can inhibit the growth of thyroid cancer cells as a result of apoptosis and/or cell-cycle arrest. Thus, it is possible that berberine could be a novel anticancer drug for the treatment of poorly differentiated or anaplastic thyroid cancer.


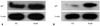
 XML Download
XML Download