

 PDF
PDF ePub
ePub Citation
Citation Print
Print


CD5(+) B cells are a subset of B cells known as regulatory B cells,1 of and are shown to negatively regulate immune responses through the production of IL-10.2 We previously described a relationship between IL-10-producing regulatory B cells and diseases such as human food allergies and atopic dermatitis.3 IL-10-producing CD19(+)CD5(+) B cell has been suggested as Br1 like Tr1, IL-10-producing T helper cell, and Br1 is associated with tolerance of food allergy. A recent study found that IL-10-producing regulatory B cells are predominantly localised within a rare CD19(+)CD5(+)CD1d(high) B cell subset.4 CD19 is a B cell marker that positively regulates B cell responses by controlling intrinsic and stimulus-dependent signalling thresholds.5 In this study, we further characterised CD5(+) B cells, based on CD19 expression levels, and found that CD19(low)CD5(+) B cells were the major subpopulation that produced IL-10.
PBMCs were obtained from venous blood of five normal subjects who had visited the Department of Allergy and Clinical Immunology at the Seoul Allergy Clinic between March and May 2010. Signed consent forms were obtained from either patients or parents. The study was approved by the Institutional Review Board of Chungnam University Hospital, Daejeon, Korea.
PBMCs were isolated from venous blood using density gradient separation and Ficoll-Hypaque (Biomedicals, Aurora, OH, USA). PBMCs were resuspended at 1×106 cells/mL in α-minimum essential medium (α-MEM; Irvine Scientific, Santa Ana, CA, USA) and stained with specific monoclonal antibodies, including allophycocyanin (APC)-labelled anti-CD19 (eBioscience, San Diego, CA, USA), phycoerythrin (PE)-Cy7-labelled anti-CD5, (eBioscience) and fluorescein (FITC)-labelled Annexin V (eBioscience). Cells were prepared at a concentration of 1×106 cells were suspended in 100 µL of FACS staining buffer (eBioscience, San Diego, CA, USA) in eppendorf tubes. Monoclonal antibodies for surface staining were added at a concentration of 1 µg/mL according to the manufacturer's instructions. The cells were incubated for 1 hour in the dark and washed three times with FACS staining buffer. The cells were then resuspended in 500 µL of FACS staining buffer prior to flow cytometric analysis.
Following staining for CD5, CD19, and Annexin V, the cells were fixed and permeabilised using a fixation/permeabilisation kit (eBioscience). After fixation/permeabilisation, the cells were stained with a PE-labelled anti-IL-10 monoclonal antibody (eBioscience). The stained cells were acquired on a FACSCaliber (BD Biosciences, Milpitas, CA, 852USA), and the data were analysed with CellQuest software (BD Biosciences).
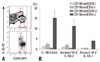
CD19 staining of PBMCs revealed two subpopulations of CD19(+) B cells, CD19(high) and CD19(low). Histogram of CD19 expression clearly, showed two peaks of CD19(+) cells (Fig. 1A). CD5 staining was used to further characterise these cells. CD19(high) and CD19(low) B cells were clearly separated by resolving with the expression of CD19 and CD5 (Fig. 1B), with SSC and CD19 expression (Fig. 1C) and with FSC and CD19 expression (Fig. 1D). Furthermore, apoptosis frequencies were 4.33% in CD19(-) non-B cell, 4.33% in CD19(high) B cells, and 16.95% in CD19(low) B cells (Fig. 2).
Among the CD5(-) cells, 22.68% were CD19(+) and 10.01% were CD19(high), whereas, 12.09% were CD19(+) and 3.59% were CD19(high) among the CD5(+) cells (Fig. 3A). Both CD5(+) and CD5(-) B cells showed CD19(high) and CD19(low) B cell subpopulations. According to the expression of CD5, CD19(low) cells showed both CD19(low) CD5(+) B cells and CD19(low)CD5(-) B cells (Fig. 3B). Based on these data, CD19(+) B cells were classified into four groups (Fig. 3C).
As shown in a representative plotting (Fig. 4), predominant production of IL-10 was observed from a CD19(low)CD5(+) B cell population. IL-10 was produced predominantly from CD19(low)CD5(+) cells among B cells (Fig. 5A). The frequencies of IL-10-producing cells were as follows: 15.2±3.4% in CD19(low)CD5(+) B cells; 0.5±0.2% in CD19(high)CD5(+) B cells; 2.3±0.7% in CD19(low)CD5(-) B cells; and 1.1±0.4% in CD19(high)CD5(-) B cells (Fig. 5B). Interestingly, most of the IL-10-producing CD19 (low)CD5(+) B cells were apoptotic. The frequencies of apoptotic IL-10-producing cells were as follows: 11.2±3.0% in CD19(low)CD5(+) B cells; 0.5±0.1% in CD19(high)CD5(+) B cells; 1.9±0.5% in CD19(low)CD5(-) B cells; and 0.7±0.2% in CD19(high)CD5(-) B cells. The frequencies of non-apoptotic IL-10-producing cells were as follows: 4.0±0.4% in CD19(low)CD5(+) B cells; 0.1±0.1% in CD19 (high)CD5(+) B cells; 0.4±0.2% in CD19(low)CD5(-) B cells; and 0.4±0.2% in CD19(high)CD5(-) B cells.
B cells are classically divided into CD5(+) B1 cells and CD5(-) B2 cells. In this study, we further subdivided these two groups of cells on the basis of CD19 expression level (Fig. 1) and found that IL-10 was mainly produced by the CD19(low)CD5(+) B cell subpopulation (Figs. 4 and 5). CD19 is a member of the immunoglobulin superfamily and is expressed only on B cells and follicular dendritic cells.6 CD19 is generally considered to be a positive regulator of both intrinsic and stimulus-dependent signalling thresholds in B cells.5 Although we found that IL-10 was predominantly expressed by the CD19(low)CD5(+) B cell subpopulation, previous reports have shown that intracellular IL-10 expression by B cells is diminished in CD19-deficient mice.7 In this study, IL-10 was found to be mainly produced by CD19(low)CD5(+) B cells however, CD19 also seemed to be necessary for IL-10 production by CD5(+) B cells.
B cells do not express IL-10 if not activated in vitro.8,9 In our recent study, IL-10 producing regulatory B cells proliferated in response to food allergens in the tolerant subjects.10 Humans are challenged continuously by tolerized antigens, including food antigens, and the present study showed that IL-10-producing regulatory B cells seem to be present in normal subjects.
CD19(+)CD5(+) regulatory B cell-mediated suppression is important for maintaining peripheral tolerance and is directly mediated by IL-10.9 CD19 overexpression allows anergic B cells to escape tolerance by converging with BCR-independent pathways.11 Our finding that IL-10 is predominantly produced by CD19(low)CD5(+) B cells is consistent with the idea that CD19 overexpression leads to a B cell subpopulation that does not produce IL-10, and therefore loss of IL-10-mediated peripheral tolerance.
CD19(high)CD5(+) and CD19(low)CD5(+) B cell subpopulations were clearly discriminated by their differential production of IL-10 and frequency of apoptosis as measured by flow cytometry (Figs. 3, 4 and 5). CD19 expression levels regulate B cell development in humans,12 and the degree of CD19 engagement correlates with B lymphocyte activation and humoral immunity in mice.13 CD81 has been shown to regulate CD19 expression in post-endoplasmic reticulum compartments, directly associate with CD19, and participate in the CD19-C21-CD81 signalling complex.14 CD19 is a highly glycosylated molecule and is expressed in two different glycoforms in B cells.15 Additional studies are needed to determine whether regulation of CD19 expression by other molecules has an effect on IL-10 production by CD5(+) B cells and also whether CD19(low)CD5(+) and CD19(high)CD5(+) B cells express differentially glycosylated forms of CD19.
CD19 amplifies B lymphocyte signal transduction by regulating Src family protein tyrosine kinases.16 Immunoregulatory roles for IL-10-producing B cells and regulatory B cells have been described in settings of inflammation, autoimmunity, and cancer.17 The functional balance between CD19 (high)CD5(+) B cells and immunosuppressive CD19(low)CD5(+) B cells may also be important for regulation of immune responses.
Several phenotypically diverse regulatory B cell subsets have been identified in recent years. CD1d(high)CD5(+) B cells,4 as well as B cell subtypes that express a CD5 isoform from alternative exon 1B (E1B),18 have been reported to produce IL-10. In patients with systemic lupus erythematosus, IL-10 production in B cells is reportedly confined to the CD154(+) subset.19 Here, we also propose that IL-10-producing CD5(+) regulatory B cells is confined to the CD19 (low)CD5(+) B cell subset.
In conclusion, division of CD5(+) B cells into CD19(high) CD5(+) and CD19(low)CD5(+) subsets revealed that IL-10 was predominantly produced by CD19 (low)CD5(+) B cells that simultaneously underwent apoptosis. The immunological role of CD19 in IL-10 production by CD5(+) B cell and the functional differences of B cell subsets expressing different levels of CD19 require further investigation.




 XML Download
XML Download