

 PDF
PDF ePub
ePub Citation
Citation Print
Print


H. pylori-associated acute and chronic antral inflammation has been associated with the expression of cyclooxygenase-2 (COX-2) in gastric epithelial cells,1 which is strongly correlated with the extent of chronic inflammatory cell infiltrate.2 Chronic expression of COX-2 is related to H. pylori-associated gastric carcinogenesis in addition to propagation of gastric inflammation since prostaglandins produced via COX-2 are reported to contribute to inflammation3 and carcinogenesis.4 Our previous study demonstrated that oxidant-sensitive transcription factor NF-κB mediates COX-2 expression, which may be related to cell proliferation in gastric epithelial cells.5 H. pylori-induced activation of NF-κB mediates the expression of several genes involved in inflammation, such as IL-86 and adhesion molecules intergrin α5, in gastric epithelial AGS cells.7 Proteinase-activated receptor 2 (PAR2) regulates cell proliferation and enhances COX-2 expression in human pancreatic cancer cells8 and integrin expression in H. pylori-infected gastric epithelial cells.9
Proteinase-activated receptors (PARs) are G protein-coupled receptors that are activated by the cleavage of their N-terminal domains by proteases.10,11 Proteinase-activated receptor 2 (PAR2) activation induces G protein-mediated signal transduction, generation of inositol triphosphate, mobilization of intracellular calcium, the activation of mitogen-activated protein kinase, cell growth, and the release of cytokines (IL-6, IL-8) and prostaglandins.12,13 PAR2 is activated by multiple trypsin-like serine proteases including trypsin and tryptase.13 Inflammatory cytokines (interleukin-1α, tumor necrosis factor-α) increase mRNA expression of PAR2.14 Invasive breast and pancreatic cancer cells express high levels of PAR2 compared with normal cells.15 These studies show the positive relationship between PAR2 expression and carcinogenesis. Previously, we demonstrated that H. pylori induced the expression and activation of PAR2 by stimulating the expression of trypsinogens and trypsin in gastric epithelial AGS cells.9 In addition, inhibition of PAR2 activation by a soyben trypsin inhibitor (SBTI) suppressed H. pylori-induced expression of integrins in gastric epithelial cells.9 Therefore, PAR2 seems to play an important role in H. pylori-associated gastric inflammation and/or carcinogenesis by mediating the induction of inflammatory and carcinogenic genes, including COX-2, in gastric epithelial cells.
In addition, the genetic differences of H. pylori isolates play a role in the clinical outcome of the infection, particularly H. pylori-virulence associated genes such as vacA, cagA, and iceA genes.16 Infection by cagA strain is more likely to result in peptic ulceration, atrophic gastritis, and gastric carcinoma.17,18 Presence of cagA or vacA in H. pylori strain showed different expression of genes as compared to cagA negative or vacA negative H. pylori in gastric epithelial AGS cells.19 The studies suggest that the presence of virulence factors (vacA, cagA, and iceA) and isotypes of each virulence factor (vacA s1b m2, vacA s1c m1, iceA1, iceA2 etc.) in H. pylori strain are important to determine disease incidence related to H. pylori infection. Since the predominant genotype of H. pylori in Korea has been reported to be cagA positive and vacA positive genotype,20 H. pylori in Korean isolates may be used to determine the pathogenic mechanism of H. pylori-induced gastric diseases in Korea.
In the present study, we investigated whether H. pylori-induced COX-2 expression is mediated by PAR2 via activation of Gi protein and Src kinase in gastric epithelial AGS cells. Thus, the cells were transfected with sense oligonucleotide (S ODN) and antisense oligonucleotide (AS ODN) for PAR2 and cultured in the presence of H. pylori. In other sets of experiments, the cells were treated with Gi protein blocker pertussis toxin, Src kinase inhibitor herbimycin A or soybean trypsin inhibitor for 1 hour and cultured in the presence of H. pylori. Subsequently, mRNA and protein expression of COX-2 were determined by RT-PCR analysis and Western blot analysis, respectively. Pertussis toxin inhibits Gi protein by coupling the inhibitory receptors to the adenylate cyclase system.21 Herbimycin A is known to be an irreversible inhibitor of Src kinase.22
We used HP99 which was isolated from gastric antral mucosa of Korean patients with gastric and duodenal ulcer, and identified it as cagA+, vacA s1b, m2, iceA1 H. pylori strain.23 H. pylori was added to human gastric epithelial AGS cells (ATCC CRL 1739, American Type Culture Collection, Manassas, Virginia, USA) at a bacterium/cell ratio of 100 : 1 in a 3 mL volume. The cells were transfected with S ODN and AS ODN for PAR2 for 16 hours and cultured in the presence of H. pylori. In other sets of experiments, the cells were treated with Gi protein blocker pertussis toxin (400 ng/mL), Src kinase inhibitor herbimycin A (10 µM), or soybean trypsin inhibitor (1, 2, 5 nM) for 1 hours and cultured in the presence of H. pylori. mRNA and protein expression of COX-2 were determined by reverse transcription-polymerase chain reaction (RT-PCR) analysis (at 12 hours) and Western blot analysis (at 24 hours), respectively. For ODN preparation, single-stranded ODNs were prepared commercially (GIBCO-BRL, New York, USA). ODNs were phosphorothioate-modified to reduce intracellular nuclease digestion. AS ODN and S ODN target the ATG start codon of the PAR2 mRNA. The sequence of PAR-2 AS ODN was 5'-TCCGCATCCTCCTGGAA-3', and that of PAR2 S ODN was 5'-TTCCAGGAGGATGC GGA-3'. For RT-PCR analysis, the primers used were: PAR2, forward 5'-GATGGCACATCCCACGTC-3', reverse 5'-GGCATGTATGTGATAGGC-3', giving a 288 bp PCR product; COX-2, forward 5'-TTCAAATGAGATTGTGGGAAAATTGCT-3', reverse 5'-AGATCATCTCTGCCTGAGTATCTT-3', giving a 296 bp PCR produc; β-actin, forward 5'-ACCAACTGGGACGACATGGAG-3', reverse 5'-GTGAGGATCTTCATGAGGTAGTC-3', giving a 354 bp PCR product. The PCR products were amplified and visualized by UV transilumination.23 For Western blot analysis, the proteins were detected with polyclonal antibodies for PAR2 (Catalog # SC-13504) and COX-2 (Catalog # SC-19999), perchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA), at 1 : 1000 dilation and the immunoreactive proteins were visualized by enhanced chemiluminescence.24
Since mRNA and protein expressions of COX-2 were evident at 12 hours and 24 hours in our previous study,24 in the following experiments using transfection with S ODN and AS ODN for PAR2 or treatment with Gi protein blocker pertussis toxin, Src kinase inhibitor herbimycin A and SBTI, respectively, the time points of 12 hours and 24 hours for mRNA and protein expressions of COX-2, respectively, were used. β-Actin (for mRNA) and actin (for protein) were constitutively expressed in the cells and not changed with culture period.
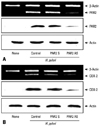
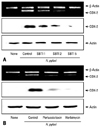
To determine direct involvement of PAR2 in the expression of COX-2, the cells were transfected with PAR2 AS ODN or S ODN and cultured in the presence of H. pylori for 12 hours (mRNA) or 24 hours (protein) (Fig. 1). Transfection efficiency of ODN was determined by observing mRNA and protein levels of PAR2 in the cells transfected (Fig. 1A). H. pylori-induced PAR2 expression was inhibited in the cells transfected with AS ODN, but not changed in those transfected with S ODN. H. pylori-induced expression of COX-2 was similarly inhibited in the cells transfected with AS ODN compared to those in the cells transfected with S ODN (Fig. 1B). Since protein level and the activity of trypsin increased in H. pylori-infected AGS cells,9 trypsin may activate PAR2 in AGS cells. Previously, we showed the increase of intracellular Ca mobilization by PAR2 activation in H. pylori-infected AGS cells,25 which was inhibited by SBTI (data not shown), demonstrating that SBTI in AGS cells suppresses PAR activation induced by H. pylori. Using SBTI, we determined whether SBTI suppresses H. pylori-induced expression of COX-2 in AGS cells. As seen in Fig. 2A, H. pylori-induced expression of COX-2 was inhibited dose-dependently by SBTI. G protein coupled receptor (GPRC) signalings have been shown to be inhibited by pertussis toxin, which inactivates ADP-ribosylation of α subunits of the Gi class in various cells,21,26 and Src family non-receptor tyrosine kinases play important roles in PAR-dependent GPCR signalings.27,28 Therefore, using Gi protein blocker pertussis toxin and Src kinase inhibitor herbimycin A, we examined the involvement of Gi protein and Src kinase in H. pylori-induced COX-2 expression in AGS cells. Fig. 2B shows that pertussis toxin and herbimycin A inhibited the expression of COX-2 in HP99-infected AGS cells. These results demonstrated that Gi protein and Src kinase are involved in PAR2-mediated COX-2 expression in H. pylori-infected gastric epithelial cells. Gi protein is known to mediate the activation of NF-κB in lung cancer cells29 and keratonocytes,30 and the connection of Src kinase to NF-κB has been reported in toll-like receptor-activated signaling31 and epidermal growth factor-induced cell proliferation.32 Since H. pylori induces the activation of NF-κB whose binding site is located in the promoter region of COX-2 gene, both Gi protein and Src kinase may be related to COX-2 expression via NF-κB pathway in addition to PAR2 pathway in H. pylori-infected gastric epithelial cells.
Recent studies demonstrate that COX-2 prevents fas-induced liver injury through up-regulation of epidermal growth factor receptor.33 COX-2 expression is not associated with the risk of cervical cancer in Korean population.34 Therefore, gastric carcinogenesis or inflammation may not be fully dependent on COX-2 expression in gastric epithelial cells or gastric mucosal tissues. Further study should be performed to determine the role of COX-2 in gastric carcinogenesis or inflammation at molecular and cellular levels. Recent study showed that advanced oxidation protein products were increased in serum of gastric cancer patients.35 However, the activities of antioxidant enzyme catalase in serums were not different between H. pylori-positive and negative patients. Since oxygen radicals are important in transcription of COX-2, the relation between oxidative stress and PAR2-mediated signaling should be investigated. The novel finding of the present study is that H. pylori in Korean isolates induced COX-2 expression, which is mediated by PAR2 through activation of Gi protein and Src kinase in gastric epithelial cells. Specific targeting of the activation of PAR2 may be beneficial for prevention or treatment of H. pylori-associated gastric inflammation and carcinogenesis in Korea.
 XML Download
XML Download