

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Elevated plasma level of native low-density lipoprotein (nLDL) contributes to numerous cardiovascular diseases including hypercholesterolemia, atherosclerosis, hypertension, heart failure, and diabetes, all of which are closely linked to vascular migration and proliferation1-3 and oxidative stress.4 It has been demonstrated that the in vitro observations of nLDL stimulation on vascular cells closely resemble many of the early changes observed both in vivo and ex vivo in human and animals during hypercholesterolemia.5-8
Reactive oxygen species (ROS) have deleterious effects on cardiovascular function.9 In nLDL-and oxidized LDL-stimulated vascular smooth muscle cells (VSMCs), O2-contributes to cell proliferation, which is particularly important to the pathophysiological conditions of in-stent restenosis, transplant vasculopathy, and vein bypass graft failure. Furthermore, pretreatment with superoxide scavengers and superoxide dismutase completely blocks the cell proliferation,10,11 implying that O2- is involved in nLDL-induced cell proliferation. On the other hand, intracellular H2O2 may mediate the mitogenic activity of growth factors through inhibition of protein tyrosine phosphatases.12 These observations suggest that ROS such as O2- and H2O2 play different physiological roles in VSMCs. Therefore, the pathophysiological role of nLDL-dependent H2O2 in VSMCs has to be elucidated.
Increased production of IL-8 as an C-X-C chemokine is involved in the recruitment of mononuclear leukocytes and stimulates the formation of atherosclerotic plaques made by macrophages, endothelial cells, and SMCs.13,14 Furthermore, VSMCs migration is associated with progression of atherosclerosis through the invasion into the intima where the plaque is formed.15 We previously reported that nLDL stimulation increased IL-8 production in human aortic SMCs (hAoSMCs) via phosphorylation of p38 MAPK and activation of transcriptional factor activator protein-1 (AP-1) with NF-kB participation. However, little information exists on nLDL-dependent signal transduction for the production of IL-8 in hAoSMCs.
Therefore, we examined the nature of the receptor and ROS responsible for nLDL-induced IL-8 production and the pathophysiological role of IL-8 played hAoSMCs migration.
MATERIALS AND METHODS
Materials
Cholera toxin, pertussis toxin, heparin, and genistein were purchased from Calbiochem. All other reagents, such as H2O2 and polyethylene glycol conjugated-catalase (PEG-Cat), were purchased from Sigma unless otherwise stated.
Cell culture
hAoSMCs were purchased from Clonetics and cultured in SmGM-2 Bullet kit medium (Clonetics) in 5% CO2 at 37℃. Twenty-four hours before experiments, growth medium was removed and replaced with DMEM medium (Gibco) containing 0.1% FBS (Gibco) for 24 hours (maintenance medium).
Isolation of n-LDL
n-LDL (density 1.019-1.063 g/mL) was isolated from the plasma of normocholesterolemic subjects (serum cholesterol <6.2 mmol/L) by differential ultracentrifugation as previously described.16 During LDL manipulation and storage, particular precautions were taken in order to maintain nLDL integrity and prevent oxidation. The content of endotoxin was below the detection limit (<1 ng/mL), as measured using an endotoxin assay kit (Pharmingen). Prepared nLDL was confirmed by thiobarbituric acid reactive substances (TBARS) assay measurement using malondialdehyde as a standard and compared its oxidation with commercial nLDL and Ox-LDL (Intracel Co., Frederick, MD, USA).
Immunofluorescence microscopy
Pretreated cells grown on gelatin-coated glass were stained with DCF-DA (2',7'-dichlorofluoroscein diacetate, 10 µmol/L), followed by incubation at 37℃ for 30 minutes. Cells were washed three times with phosphate-buffered saline (PBS) and imaged at 488/535 nm using an epifluorescence microscope (Olympus BX51).
Determination of IL-8 concentration
The amounts of IL-8 released into the media were assayed using a human IL-8 ELISA Kit II (OptEIA, BD Biosciences, NJ, USA) according to the manufacturer's instructions.
Western blot analysis
Cells treated with different chemicals were lysed in SDS sample buffer (62.5 mmol/L Tris pH6.8, 2% SDS, and 10% glycerol). Each sample was resolved on a 10% SDS-PAGE, transferred onto PVDF membranes (Pall), analyzed with antibodies according to the supplier's instructions, and then visualized with peroxidase and an enhanced-chemiluminescence system (ECL kit, Amersham, NJ, USA). Polyclonal phospho-p38 MAPK was purchased from Cell Signaling Technology and normalization was performed with the smooth muscle cell (SMC)-specific α-actin antibody (BD Biosciences, NJ, USA).
Migration assay
Transwell plate (13 mm gelatin-coated polycarbonate filter, 8 µm pore size, Costar Corp.) was used for the migration assay. Briefly, suspended cells (1×105) were added to the upper chamber of the Transwell insert, and IL-8 or neutralizing antiserum against IL-8 (R&D system, MN, USA) was mixed with the medium [DMEM+1% FBS+0.1% bovine serum albumin (BSA)] in the lower chamber. After the transwell was incubated for 8 hours, the filters were fixed with methanol (10 minutes at 4℃) and stained with hematoxylin. The number of migrated cells was determined by microscopy (Olympus BX51, 400X high power field).
RESULTS
nLDL increases IL-8 production through Gαi/o-coupled receptor in hAoSMC
First, we tested whether prepared LDL underwent any change such as oxidative modification. However, LDLs prepared in 3 independent experiments showed no oxidation, because the value was -0.60±0.06 µmol/L in TBARS assay. This value was comparable to commercially available nLDL, which was -0.55±0.04 µmol/L, whereas, oxidized LDL was 7.95±0.60 µmol/L.
Since we previously demonstrated that stimulation of hAoSMCs with nLDL increased IL-8 production via p38 MAPK activation 16, we attempted to determine the nature of the receptor responsible for IL-8 production. As shown in Fig. 1A, increased IL-8 production by nLDL from 4.96±0.36 ng/mL to 14.03±0.56 ng/mL (Fig. 1A, *vs. untreated, p<0.01) was completely blocked by pretreatment of the cells with PTX (Fig 1A, **vs. nLDL=3.23±0.03 ng/mL vs. 14.03±0.56 ng/mL, p<0.01). This effect of pertussis toxin (PTX) was associated with inhibition of p38 MAPK phosphorylation (Fig. 1B). On the other hand, cholera toxin (CTX) treatment had no effect on IL-8 production (Fig. 1A, #vs. nLDL=13.63±0.39 ng/mL vs. 14.03±0.56 ng/mL, not significant) and p38 MAPK phosphorylation (Fig. 1B). Furthermore, inhibitors related to receptor-associated signal transduction, such as heparin for the classical nLDL receptor or genistein for receptor tyrosine kinase, were employed, however, these agents had no inhibitory effect on IL-8 production (Fig. 1A, nLDL+heparin vs. nLDL=13.91±0.84 ng/mL vs. 14.09±1.38 ng/mL, not significant; nLDL+genistein vs. nLDL=13.89±0.40 ng/mL vs. 14.09±1.38 ng/mL, not significant) and p38 MAPK activation (Fig. 1C). In addition to nLDL, sphingosine-1-phosphate (S1P) and lysophosphatidic acid (LPA) have been known as agonists binding to Gi-protein coupled receptors. Therefore, nLDL was compared with S1P and LPA in terms of IL-8 production. Unlike nLDL, however, Fig. 1D shown that S1P and LPA did not induce IL-8 production (*vs untreated=13.57±0.29 ng/mL vs. 5.53±0.59 ng/mL, p<0.01; S1P or LPA vs. untreated=5.56±0.34 ng/mL (S1P) or 5.51±0.50 ng/mL (LPA) vs. 5.53±0.59 ng/mL, not significant).
PTX-sensitive G-protein coupled receptor mediates intracellular H2O2 generation in nLDL-stimulated hAoSMC
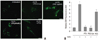
We next tested whether nLDL induces H2O2 generation and also H2O2 generation is dependent on PTX-sensitive G-protein coupled receptor. As shown in Fig 2, incubation of hAoSMCs with nLDL resulted in a significant increase of DCF fluorescence (*vs. untreated=7.42±0.72 vs.1.00±0.14, p<0.01, n=3). On the other hand, preincubation of the cells with PTX (#vs. nLDL=1.10±0.10 vs. 7.42±0.72, p<0.01, n=3) or PEG-Cat (##vs. nLDL=0.92±0.19 vs. 7.42±0.72, p<0.01, n=3) significantly prevented the increase of nLDL-induced DCF fluorescence. Furthermore, H2O2 markedly increased DCF fluorescence (**vs. untreated=5.36±0.38 vs. 1±0.14, p<0.01, n=3). Fig. 2B shows relative fluorescence values as bar graph from 3 independent experiments.
Increased intracellular H2O2 plays a key role in nLDL-dependent p38 MAPK phosphorylation and IL-8 production
Since p38 MAPK is a critical component of redox-sensitive signaling pathways in VSMCs,17-19 we tested whether nLDL-induced p38 MAPK activation and IL-8 production were associated with increased intracellular H2O2 generation. As shown in Fig. 3A, nLDL-induced p38 MAPK activation was completely blocked with pretreatment of the cells with cell-permeable PEG-Cat. Furthermore, increased IL-8 production by nLDL was also abrogated by incubation with PEG-Cat (Fig. 3B, *vs. untreated=15.12±0.36 ng/mL vs. 4.83±0.29 ng/mL, p<0.01; #vs. nLDL=4.98±0.41 ng/mL vs.15.12±0.36 ng/mL, p<0.01). Interestingly, exogenous H2O2 at a low concentration of 10 µmol/L induced the phosphorylation of p38 MAPK as early as 10 minutes (Fig. 3C) and increased IL-8 production in a dose-dependent manner (Fig. 3D, *vs. untreated control=6.42±0.39 ng/mL vs. 4.95±0.36 ng/mL, p<0.05; #vs. untreated=10.23±1.26 ng/mL vs. 4.95±0.36 ng/mL, p<0.01). Since exogenous H2O2 at concentrations higher than 20 µmol/L significantly decreased cell viability as early as 1 hour, we failed to completely mimic IL-8 production to the level of nLDL stimulation. Therefore, these data suggest that intracellular localization of H2O2 generation could be important in the activation of redox-sensitive protein kinases.
IL-8 increases the migration of hAoSMC
We next examined whether the secreted IL-8 had any effect on the migration of hAoSMCs that is one of critical phenomena of hAoSMC during atherogenesis. Therefore, the effect of IL-8 on hAoSMC migration was determined by Transwell assay as described in the methods section. As shown in Fig. 4, IL-8 treatment at 15 ng/mL significantly increased hAoSMCs migration (*vs. untreated=1.88±0.30 vs. 1.04±0.30, p<0.01). The migration activity of IL-8 was completely blocked with neutralizing antiserum (0.5 µg/mL) against IL-8 (#vs. IL-8=0.98±0.16 vs.1.88±0.30, p<0.01). PDGF-BB as a positive control increased the migration as much as 2.45±0.11 fold.
DISCUSSION
Herein, we have demonstrated that nLDL stimulation transduces a signal via the PTX-sensitive G-protein coupled receptor and redox-imbalance by generating H2O2, both of which are important signaling events in p38 MAPK activation and IL-8 production in hAoSMCs. Furthermore, IL-8 at a concentration of 15 ng/mL significantly increased hAoSMCs migration that was blocked with IL-8-neutralizing antiserum.
nLDL evoked proinflammation by upregulating IL-8 production via phosphorylation of p38 MAPK and activation of transcription factor, AP-1,16 and also induced proliferation via PTX-sensitive G-protein coupled receptor-dependent protein kinase C (PKC) activation in hAoSMCs.20 Consistent with the effect of nLDL on hAoSMC proliferation, increased IL-8 production was completely blocked by PTX treatment. However, CTX, Gαs-protein coupled receptor inhibitor, heparin (classical LDL receptor inhibitor), and genistein (a tyrosine kinase inhibitor), had no effect on nLDL-induced IL-8 production (Fig. 1). These data suggest that nLDL transfers signals for IL-8 production through the common Gαi/o-protein coupled receptor instead of both the conventional LDL receptor, LDLR, and endothelial differentiation gene (EDG) receptor. The Gαi/o-protein coupled receptor appears to be responsible for activation of Erk1/2 MAPK20 and p38 MAPK during nLDL signal transduction.
Since that atherogenesis is generally viewed as a chronic inflammatory disease, an altered redox-state results in enhancing the expression of proinflammatory genes that facilitate interactions between endothelium and leukocytes. Increased expression of proinflammatory genes is mediated through redox-sensitive transcription factors, such as NF-kB, AP-1, Egr-1, and HIF-1β, which are also involved in growth, vascular remodeling, and atherogenesis of VSMCs. ROS exerts intracellular signaling processes by activating mitogen activated protein kinase (MAPK) and receptor- or non-receptor-tyrosine kinases, although previous studies have revealed contradictory results on the physiological roles of intracellular H2O2 and O2- in VSMCs. VSMCs proliferation induced by O2- generating LY8358321 and nLDL-dependent proliferation is blunted by the administration of PEG-SOD,21,22 suggesting that intracellular O2- plays an important role in VSMCs proliferation. On the other hand, angiotensin II-dependent p38 MAPK activation is a process associated with H2O2 generation19 and activated p38 MAPK stimulates cell migration.23 These results are consistent with our finding that nLDL increased intracellular H2O2 generation, resulting in activation of p38 MAPK associated with IL-8 production (Fig. 3).
Every cellular component in the vascular wall is known to be a potential source of IL-8.16,24,25 Produced IL-8 has some functional characteristics similar to other cytokines. IL-8 is resistant to temperature, proteolysis, and acidic environments, making it as an ideal candidate molecule for acute inflammation.26 Another characteristic of IL-8 is its relative longevity at sites of acute inflammation.26 IL-8 is produced early in the inflammation response, but remains active for a prolonged period of time, even days and weeks. A third interesting aspect of IL-8 is that the expression is highly sensitive to oxidants and anti-oxidants substantially reduce IL-8 gene expression.27 Furthermore, IL-8 contributes to atherogenesis by modulating monocyte-endothelial interaction,13 stimulating VSMCs proliferation,15 inhibiting tissue inhibitor of metallopeptidase (TIMP-1) expression,28 and inducing angiogenic activity.29 On the other hand, the neutralization of IL-8 significantly reduces the degree of necrosis in a rabbit model of myocardial ischemia-reperfusion injury.30 The present study together with all the above-mentioned observations indicate that IL-8 may be either a marker or a potential therapeutic target of atherosclerosis.31
In summary, this study demonstrated that nLDL transferred a signal through PTX-sensitive receptor and increased H2O2 generation that were key phenomena for p38 MAPK activation and IL-8 production in hAoSMCs. Furthermore, IL-8 treatment increased migration activity of hAoSMCs.



 XML Download
XML Download