

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Brain-derived neurotrophic factor (BDNF) is directly involved in neurite outgrowth and regulates the survival, differentiation, and maintenance of function in different neuronal populations.1 Tyrosine kinase-coupled receptor (TrkB) is the primary signal transduction receptor for BDNF. BDNF is the central neurotrophic factor (NTF) for neurons and neurogliocytes, and plays an important role in hippocampal aging. In fact, BDNF and TrkB expression have been shown to decrease both in the normal aging hippocampus and in the Alzheimer's patient hippocampus, while they significantly increase during learning-related events.2-5
The hippocampus is one of the important brain areas that is connected with learning and memory, and it is also an area that is very susceptible to stress and senescence.6,7 It is well-established that stress is a conspicuous factor of neuronal injury and can trigger degenerative cellular processes in the limbic system.8,9 Single or repeated immobilization stress treatments have been reported to decrease BDNF mRNA throughout the hippocampus.10-12 In contrast, BDNF mRNA expression measured by in situ hybridization increased as early as 15 min in most hippocampal regions and was significantly augmented after 180 min of stress exposure. Adlard reported that the expression of BDNF obviously decreased at time-course 5 and 10 hour after binding stress, but the expression of TrkB mRNA showed no change in dentate gyrus and hippocampus after single stress.9
Although it is recognized that BDNF is the most important NTF in critical CNS functions, including neuronal development and differentiation, time-course studies of different stress applications have not yet been investigated in terms of aging responses. In the present study, we investigated whether both acute stress (AS) and repeated chronic mild stress applications might modify expression of hippocampal BDNF mRNA and protein and TrkB mRNA content, or whether there was an age-related change in BDNF expression in the hippocampus of rats.
Animal behaviors including attacking behavior, languishing behavior, and excreting behavior evidently change in stressful conditions. Aging is an important factor affecting the behavior of animals and human being. The exploratory behaviors and locomotor activities during open field (OF) test can reveal the rat's adaptation to a new environment,13,14 which indicates the cognitive ability of rats.
Therefore, in order to clarify the changes in BDNF after different stresses and the effect of aging on the expression of BDNF protein, we studied the expression of BDNF mRNA in the hippocampus following forced-swimming stress by reverse transcription-polymerase chain reaction (RT-PCR) with an endogenous internal standard, and detected the expression of BDNF protein by Western blotting.
MATERIALS AND METHODS
Animals
Male Sprague-Dawley rats aged 2 months and 22 months were obtained from the Animal Experiment Center of Binzhou Medical University, and housed for 1 week prior to the experiment under a constant temperature (21 ± 1℃) and lighting regimen (light on from 7:00 am to 7:00 pm). Food pellets and water were available ad libitum throughout the experiments. Young and aged rats were randomly divided into three subgroups: the AS group (n = 30), the chronic mild repeated stress (CMRS) group (n = 30), and a control group (n = 8).
Stress models
AS model: animals were forced to constantly swim in 4℃ water in a 70×40×80 cm aquarium (35 cm depth of water) for 10 minutes on the day of the experiment (Fig. 1A). CMRS model: animals were forced to swim in 25℃ water for 10 minutes at 8:30 am, which was enforced for 21 consecutive days in the same aquarium (Fig. 1B). After the conclusion of the different stress tests, animals were immediately decapitated. In the control group, unstressed animals were handled daily, and on the day of the experiment, they were sacrificed at the same time as the stressed animals.
Open field (OF) test
The OF box was a 90×90×45 cm wooden box used to study the cognitive and emotional reaction by observing the animal's behaviors.15 Its bottom was divided into 5×5 cm squares; the square in the middle was designated the center square, the others were peripheral squares. For the stress test, eight rats were selected randomly from the two age groups used for the CMRS test. The OF test was performed on day 1, 2, 3, 4, 5, 10, 15, and 21, immediately following the CMRS test. Each animal was placed in the center of the OF box and observed for 3 minutes. The indices of the test included the number of square crossings, number of grooming events, time spent on the center square, vertical movement scores, and number of stools produced. All indices were observed and recorded by two individuals who did not know the purpose of this test. The OF box was cleaned after each test session.
Corticosterone radioimmunoassay
To determine the efficacy of different times of stress application, plasma corticosterone was analyzed. Before the rats were killed, blood samples were collected by intracardiac puncture and were centrifuged at 4℃ to separate the plasma, which was stored at - 20℃ until assayed for corticosterone. Plasma corticosterone levels were measured by a radioimmunoassay kit (ICN Biomedicals, Costa Mesa, CA, USA). The intra-assay variability of the RIA ranged between 3.1 and 4.5%. The sensitivity of the assay was 5.7 ng/mL.
Western blotting
Animals of young and aged groups were anesthetized with sodium pentobarbital (100 mg/kg, i.p.) and sacrificed immediately by decapitation at each point of 15, 30, 60, 180, and 720 min after stress, and control rats were killed under the same condition (0 min, basic expression). Their brains were removed for isolating the hippocampal tissues. For each rat, 50-100 mg hippocampal tissue was abraded and lysed in 1 mL tissue and cell lysis solution,16 then centrifuged at 15,000 g for 15 min at 4℃. The protein content of the supernatant was determined using the Bradford assay,16 then an equal amount of protein from each sample was subjected to 15% SDS-PAGE. The separated proteins were transferred to a nitrocellulose membrane and detected using an Enhanced Chemiluminescent Method (ECM) of Western Blotting system. An immunoblotting reaction was performed with a rabbit anti-BDNF polyclonal antibody (1 : 400 dilution in 1×TBS-T, and the total volume was 4,000 µL) overnight at 4℃; horseradish peroxidase (HRP-labeled goat anti-rabbit IgG) was used as the secondary antibody (1 : 10,000 dilution in 1×TBS-T, and the total volume was 5,000 µL). The expression of BDNF was determined by calculating the density ratio of BDNF to a β-actin band (the software of Image J was used to analyze the density ratio and the software was downloaded from http://rsb.info.nih.gov/ij/download.html). Anti-BDNF antibody, anti-β-actin antibody, and the secondary antibody were purchased from Wuhan Boster Company (http://www.boster.com.cn/).
RT-PCR assay
Hippocampal tissues were obtained at a total of 6 time points from young and aged groups. Total mRNA was extracted from 50-100 mg hippocampus according to the instructions of the TRIzol kit (Invitrogen).The RT-PCR kit was purchased from Promega. The primers for BDNF and TrkB were synthesized by Sbsgene Company, and the sequences of the primers were as follows:
5'AGTGATGACCATCCTTTTCCTTAC3'plus
5'CCTCAAATGTGTCATCCAAGGA3'(196bp, for BDNF)
and
5'GGCCAAGAATGAATATGGTAA 3'plus
5'TTGAGCTGGCTGTTGGTGAT 3'(485bp, for trkB)
and
5'CACAGCTAGAGGGAAATCG3'plus 5'CACCAGAGTAGTTGCGCTC 3'(348bp, for β-actin).
Reactions were performed in a 50 µL volume containing 1 µL AMV reverse transcriptase, 1 µL TflDNA polymerase, 2 µL 25 mM MgSO4, 2 µL primers, 1 µL dNTP Mix, 10 µL 5×Reaction Buffer, and 31 µL Nuclease-Free Water. The reaction sequence was 45℃ for 45 min (1 cycle), 94℃ for 2 min (1 cycle), 94℃ for 30 sec, 55℃ for 1 min, 68℃ for 2 min (30 cycles), 68℃ for 7 min (1 cycle), then a 4℃ soak. PCR products were analyzed by electrophoresis and the density ratio of target genes to the β-actin band was used to determine the levels of expression.
Statistical analysis
SPSS 11.0 software (SPSS Inc, Chicago, IL, USA) was used for calculations in this study. Mean and SEM were calculated from 6 animals per group for BDNF and TrkB mRNA. The quantitative data were expressed as Mean ± SD and statistical analysis was performed by t-tests between individual groups. One-way ANOVA was used to analyze the different expression of BDNF mRNA between young and aged groups of rats.
RESULTS
Changes in OF test indices in different CMS models
Fig. 2 shows that there were no significant differences in exploratory behaviors among handled and control animals in either age group prior to imposition of stress. With prolongation of the experimental days, the quadrant crossing, grooming, and vertical movement scores of young and aged CMRS group animals exhibited a downward trend, and the time spent in the center square clearly increased. The OF indices of the aged CMRS group showed significant decreases compared with the young group, and displayed depression-like behavior and weak stress resilience (Figs. 2 and 3).
Changes in plasma corticosterone levels with duration of stress exposure
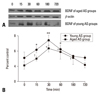
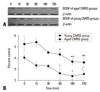
There was a large, rapid, and significant increase in plasma corticosterone levels after 15 min of AS, both in the young and the aged groups (294 ± 27 ng/mL vs. 21 ± 7 ng/mL in young control group, t = 42.89, p < 0.001; 197 ± 31 ng/mL vs. 9 ± 2 ng/mL in aged control group, t = 13.36, p < 0.001), and the highest peak of corticosterone present, respectively, at 30 min in young group and 60 min in aged groups after stress (466 ± 43 ng/mL; 328 ± 22 ng/mL), which all maintained high levels until 180 min after stress (Fig. 4). The levels of plasma corticosterone in the two CMRS groups also showed increases after stress, and reached maximal concentrations 30 min after stress (193 ± 38 ng/mL in young group; 104 ± 20 ng/mL in aged group), which were lower than those of AS groups (Fyoung = 26.58, p < 0.01; Faged = 25.46, p < 0.01) (Fig. 5).
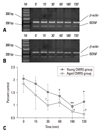
Time course expression of BDNF in the whole hippocampus after different stresses as detected by Western blotting
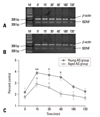
Western blotting was performed in the hippocampus to examine discrete modifications of the BDNF protein on control groups (0 min) and stress-exposed groups at 15, 30, 60, 180, and 720 min after stress. After AS, we observed a rapid increase in BDNF at 15 min (as well as 30 and 60 min) throughout the entire hippocampus, in both young and aged groups. A decrease occurred at 180 min and the levels had approached the original condition by 720 min. The maximum expression of BDNF in both young and aged AS groups were observed at 30 min (Fig. 6). After chronic repeated stress, the expression of BDNF showed a dynamic change at 15, 30, 60, 180, and 720 min, whereas the levels were lower than those measured in both the young and aged control groups. However, we observed that the BDNF protein level detected in the young group, 15 min after the last repeated stimulus, was higher than that measured at all other time points, and displayed a remarkable change from 15 min to 180 min. In contrast, the expression level in the aged group was lower than that of the young group at several time points and showed a similarly low level from 60 min to 720 min (Fig. 7).
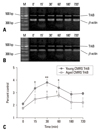
Effect of stress duration on BDNF mRNA and TrkB mRNA expression as determined by PT-PCR
To evaluate variations in the expression of BDNF and TrkB transcripts after different stresses, these were analyzed at 15, 30, 60, 180, and 720 min after stress by RT-PCR. AS application resulted in rapid up-regulation of BDNF mRNA, with a maximum expression occurring after 15 min of stress in both young and aged groups. There was a significant decrease in expression at 60 min in the aged group, but not in the young group (Fig. 8). No quantitative changes in TrkB mRNA were observed after short periods of AS, although a slight increase was seen at 30 min (Fig. 9). In contrast, following chronic stress, BDNF mRNA measured at five points exhibited decreased expression compared with those of the control groups, but the TrkB mRNA was significantly augmented in the young group during the time course after stress (Figs. 10 and 11). The levels of BDNF and TrkB transcript expression in aged rats were lower than those measured in the initial condition in aged control animals. The levels of BDNF mRNA dec ± reased after chronic stress, whereas TrkB mRNA had increased.
DISCUSSION
Animal behaviors clearly change in response to stressful conditions.17 Forced-swimming, a relatively mild physiological stress that causes no body damage, was used to build the current animal model. An OF test was used to evaluate rat behavioral changes following exposure to chronic stress. Quadrant crossing is a locomotor activity that shows the rat's movements in an open field, while grooming is an emotional response that indicates the tension of the rats in a new environment. These indices help us to evaluate the behavioral changes of rats in response to stress.18,19
In the current study, two factors led to the behavioral disturbances, namely, aging and the duration of stress. We observed that both young and aged CMRS groups displayed a decrease in square crossing, grooming behavior, and vertical movements, as well as an increase in stool production and increased retention time in the middle square compared with control groups after stress, which represented an inhibition of activities and an increase in emotional disorders. Furthermore, the behavioral activity of aged animals was noticeably weaker than that of young ones in several aspects, representing an obviously abepithymia state. Animals showed little interest in, and weak adaptability to, new surroundings under a chronic stress state. Moreover, behaviors of aged animals were fewer in number than those observed in the young group, reflecting a weaker tolerance to stress. Stress had effect on the function of pre-frontal cortex and limbic system, which were closely bound up with cognition and behavioral response.20 The level of plasma corticosterone in aged animals obviously rose, which acted on the down-regulation of the hippocampal glucocorticoid (GC) receptor synthesis and receptor binding,21,22 and the disorder of negative feedback function of hypothalamo-pituitary-adrenal (HPA) axis.22,23 Thus, senescence led to the enhanced sensitivity of animals to stress, which may result in a serious stress injury and changes of behaviors.
The limbic-hypothalamopituitary-adrenal (LHPA) axis plays an important role in stress and is the ultimate nerve pathway operating during stress responses. Corticotropin-releasing hormone (CRH), adrenocorticotropic hormone (ACTH), and GCs are important hormones in the HPA axis. GCs are the key hormones involved in stress adaptation because of their influence on multiple organs.24 GCs are known to exert feedback regulation on the functioning of the HPA axis, and can increase the release of excitatory amino acids.25 This leads to excitability toxicity of hippocampus, which is characterized by abnormal form of pyramidal neurons, neuronal atrophy, and significant decreases in cell number.26 Therefore, plasma corticosterone expression can be viewed as a symptom of stress injury during exposure to a stressor. Previous studies have indicated that 3 h incubation of hippocampal neurons with GCs depresses activity-dependent expression of BDNF mRNA.27,28 Similarly, Lauterborn, et al. reported that adrenal hormones inhibit in vivo BDNF expression.29 In contrast, Marmigère, et al.30 demonstrated that plasma corticosterone levels were strongly increased after 15, 60, and 180 min of stressor exposure, and were accompanied by a high expression of BDNF.31 In the present report, corticosterone levels in peripheral blood detected rapidly increased in response to AS at 15, 30, and 180 min in both young and aged groups, indicating an intensive stress state. Plasma corticosterone levels determined in the chronic groups also showed increases at 15 and 30 min after stress, but these levels declined to control levels over time, affirming the stress efficacy. Senescence and stress both resulted in the increase of the corticosterone,32,33 which led to the hyperfunction of hippocampus and functional deficit of learning, memory, and cognition. Meanwhile, the hippocampal neuronatrophy that was induced by sustained high concentration of GCs had an effect on the decrease of BDNF mRNA expression.34,35
BDNF is a NTF involved in critical CNS function, as well as synaptic transmission and plasticity, and it plays an important role in the survival, maintenance, and growth of neurons.36-38 The results of the present study demonstrate that AS induced a rapid increase in the expression of BDNF, BDNF mRNA, and TrkB mRNA in the hippocampus, in both young and aged animals. Rapid variations in BDNF protein induced by AS were observed at 15, 30, and 60 min after stress, which was followed by a downturn to the initial condition by 180 min. Overall, the expression of BDNF in the aged group was significantly less than that determined in the young group at 15 and 30 min. Increased BDNF mRNA levels have been previously reported, after 60 min immobilization stress, and decreased levels after 2 h stress, in the dentate gyrus, the CA3 area, and the entire hypothalamus.17,39,40 However, the expression of TrkB mRNA was not influenced by acute immobilization,35 which is contrary to the observations in the present study. In contrast, chronic repeated stress led to a notable increase in TrkB mRNA, but an obvious decrease in BDNF mRNA and proteins in the previous work.17
To fully confirm the specificity of the early increase in BDNF detected in the present report, BNDF mRNA and TrkB mRNA were measured by RT-PCR. The mRNA results confirmed that the augmentation of BDNF mRNA had already occurred 15 min after stress. No substantial increase in TrkB mRNA was seen, although there was a slightly enhanced expression compared with basic condition.
BDNF and its receptor TrkB play important roles during stress injury. After a series of acute injuries, such as cerebral ischemia, epilepsy, or cerebral trauma, the upregulation of BDNF mRNA, as well as the augmentation of TrkB mRNA, were observed in the cerebral cortex and hippocampus.31,41 Similarly, Fujhara, et al.42 demonstrated that the levels of BDNF protein and mRNA increased significantly in the hippocampus of rats that had been treated with short-term sleep-disturbance.43 Moreover, increased BDNF mRNA and protein levels occurred in the pituitary glands of rats stressed for 60 min, while decreased levels occurred following stress for 180 or 300 min.42 Scaccianoce, et al. and Smith, et al. demonstrated by RT-PCR and in situ hybridization that AS induced down-regulation of BDNF mRNA.35,44 Most researchers now view long-term and chronically repeated stress as making significant contributions to decreased expression of BDNF, as well as increased expression of TrkB mRNA. The decreased expressions of BDNF mRNA and protein were observed in the CA3 and the dentate gyrus of the hippocampus after repeated immobilization stress,45,46 together with increased expression of TrkB mRNA.
In the present study, the expression of BDNF protein and mRNA showed a significant decrease at 60, 180, and 720 min after CMRS. The levels of BNDF mRNA detected in both young and aged CMRS groups were lower than those of acute groups at several time points after stress. The expression levels of BDNF and TrkB determined in the aged groups were lower than those measured in the young group at several time points, as analyzed by Western blotting and RT-PCR. Thus, although not definitive, the results suggested that BDNF and its receptor were influenced by both the stress paradigm and by senescence. The up-regulation of TrkB mRNA measured in the CMRS groups may be a compensatory adaptation to repeated stress, similar to the change of BDNF detected in the AS groups.
We now know that unexpected and AS tends to provide an excited organism with a degree of protection for a short time. This suggests that the augmented expression of BDNF mRNA and protein might, either directly or indirectly, contribute to stress protection. However, with prolongation of the stress, if the excitation is not attenuated, secondary neuronal damage would occur.47 Thus, a second response that leads to a decrease in BDNF mRNA and protein expression occurs. The observed up-regulation of TrkB mRNA might be a further compensatory adaptation to prolonged stress-induced down-regulation of BDNF. Up-regulation of TrkB could possibly make the neurons of the hippocampus more responsive to lower levels of BNDF than are induced by chronic stress.44,46 The present study showed that the expression of BDNF and TrkB were associated with aging, which is known to attenuate mammalian stress coping capacity.48-50 The expression of BDNF and TrkB has been reported to increase during learning-related events and to decrease in the hippocampus of alzheimer's disease (AD) patients, suggesting that these are involved in learning, emotion, and age-related memory deficits. Although Lapchak, et al. determined that age did not change the prevalence or regional distribution patterns of BDNF or TrkB mRNA in the hippocampal formation throughout the lifespan of male rats,51-53 more research has shown that significant decreases with age can be detected for BDNF mRNA and TrkB mRNA in many areas of the brain.2,51,54
We compared the dynamic change of BDNF and TrkB expression in the hippocampus of young and aged rats after different stress applications in order to illustrate the effect of senescence on neurotrophin expression. A significant increase in BDNF mRNA and protein occurred both in young and aged rats during the time course after AS, accompanied by a slight change in TrkB. The levels of the aged group were generally lower than were those of the young group. Similarly, the dynamic change of BDNF and TrkB expression that was detected in chronically stressed groups was slighter than that of AS groups, and there was a distinct reduction in response in the aged group compared to the young group. Hock and Hosinger suggested that the induction of BDNF and TrkB mRNA detected in the hippocampus of AD was substantially lower than that seen in normal adult cellular tissues.55,56 More importantly, the spread of BDNF in the AD hippocampus was similar to that in naturally aging animals, suggesting that a low content of BDNF in the brain was one of the reasons for cognitive disorders and weak responsiveness to stress. Thus, we also concluded that aging could reduce the expression level of BDNF and TrkB mRNA, which might contribute to poor stress protection seen in the present study in aged rats.57
In conclusion, alterations of BDNF and its receptor of TrkB were influenced by several factors, such as a stress paradigm, stress duration, and aging. The rapid increase in BDNF mRNA, protein, and TrkB mRNA in response to AS may be part of a neuronal protective response. The decreased expression measured in aged animals exposed to chronic stress may indicate a natural loss of this protective function during senescence. Clearly, further study will be necessary to confirm the expression of BDNF and its high affinity receptors and to explore the mechanisms in up/down-regulation of BDNF expression in different brain areas, in order to further understand the complexities of neuronal responses to stress.





 XML Download
XML Download