

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Papillary thyroid cancer (PTC) is the most common thyroid malignancy.1 The biological behavior of PTC varies widely, from indolent micro-carcinomas to invasive tumors that can metastasize. Although the etiology of PTC is not well defined, a strong association with exposure to ionizing radiation exists in a minority of cases.2 Many attempts have been made to identify markers for thyroid carcinoma that can distinguish between benign and malignant lesions. However, most of them were proven to be not specific enough, including Galectin-3, BRAF, Met, hTERT, and HBME-1.3-7
Techniques designed to identify differentially expressed genes (DEGs) in tumors have become pivotal in modern pathology. Genefishing technique™ using the annealing control primer (ACP) system has recently been developed by Hwang et al. to screen for DEG transcripts.8 It uses primers that anneal specifically to the template and allows only genuine products to be amplified, thus eliminating false-positive results.
We adapted the ACP system for the identification of DEGs involved in PTCs. Differential display was performed by using kits which incorporated an ACP. We confirmed the expression patterns of 10 DEGs, and quantified their expression by RT-PCR. These results suggest that this new PCR-based technique is a very useful tool for identification of novel genes differentially expressed in PTCs.
MATERIALS AND METHODS
Patient population
Twenty-one patients diagnosed with PTC were included in this study. The frozen tissues from patients were used for RNA isolation. And the frozen tissue from only one subject was used for the differential display analysis.
All PTCs were analyzed according to various clinical-pathological parameters. Clinical staging of PTC was classified according to the Tumor Node Metastasis (TNM) classification of the International Union Against Cancer (UICC).
GeneFishing™ reverse transcription-polymerase chain reaction
First-strand cDNA synthesis
Total RNAs extracted from frozen samples were used for the synthesis of first-strand cDNAs by reverse transcriptase. Reverse transcription was performed for 1.5 hours at 42℃ in a final reaction volume of 20 µL containing 3 µg of the purified total RNA, 4 µL of 5' reaction buffer (Promega, Madison, WI, USA), 5 µL of dNTPs (2 mM each), 2 µL of 10 µM dT-ACP1 (5'-CGTGAATGCTGCGA CTACGATIIIIIT(18)-3'), 0.5 µL of RNasin® RNase Inhibitor (40 U/µL; Promega), and 1 µL of Moloney murine leukemia virus reverse transcriptase (200 U/µL; Promega). First-strand cDNAs were diluted by the addition of 80 µL of ultra-purified water for the GeneFishing™ PCR and stored at -20℃ until use.
ACP (Annealing Control Primer)-based GeneFishing™ PCR
DEGs were screened by ACP-based PCR method using the GeneFishingTM DEG kits (Seegene, Seoul, South Korea). Briefly, second-strand cDNA was synthesized at 50℃ during one cycle of first-stage PCR in a final reaction volume of 20 µL containing 3 - 5 µL (about 50 ng) of diluted first-strand cDNA, 1 µL of dT-ACP2 (10 µM), 1 µL of 10 µM arbitrary ACP, and 10 µL of 2 × Master Mix (Seegene, Seoul, South Korea). The PCR protocol for second-strand synthesis was one cycle at 94℃ for 1 minute, followed by 50℃ for 3 minutes, and 72℃ for 1 minute. After second-strand DNA synthesis was completed, the second-stage PCR amplification protocol was 40 cycles of 94℃ for 40 seconds, followed by 65℃ for 40 seconds, 72℃ for 40 seconds and 5 minutes final extension at 72℃. The amplified PCR products were separated in 2% agarose gel and stained with ethidium bromide.
Cloning and sequencing
The differentially expressed bands were extracted from the gel by using the GENCLEAN® II Kit (Q-BIO gene, Carlsbad, CA, USA), and directly cloned into a TOPO TA cloning vector (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. The cloned plasmids were sequenced with ABI PRISM® 3100 Genetic Analyzer (Applied Biosystems, Foster City, CA, USA).
The DNA sequence of each gene was confirmed by comparison with sequences in GenBank (NIH, MD, USA). The primer sequence of each gene was designed using the PRIMER 3 program (MIT, Boston, MA, USA). The synthesis of primers was performed by Bioneer Company.
RT-PCR confirmation
The differential expression of DEGs was confirmed by RT-PCR using each gene specific primer pair. The first-strand cDNA was normalized by the human beta-actin gene. The normalized cDNA was used as a template. The PCR reaction was conducted in a final reaction volume of 20 µL containing 2 - 4 µL (about 50 ng) of diluted first-strand cDNA, 1 µL of primer 5 (10 µM), 1 µL of primer 3 (10 µM), and 10 µL of 2 × Master Mix (Seegene, Seoul, South Korea). The PCR amplification protocol was an initial 3 minutes denaturation at 94℃, followed by 20 - 25 cycles of 94℃ for 40 seconds, 60℃ for 40 seconds, 72℃ for 40 seconds, and a 5 minutes final extension at 72℃. The amplified PCR products were separated in 2% agarose gel and stained with ethidium bromide.
RESULTS
Clinicopathologic features
The ages of 21 patients (2 men and 19 women) ranged between 18 and 66 years (mean age, 46 years). Clinical staging according to the TNM classification showed stage I in 3, stage II in 9, stage III in 8, and stage IVA in 1 patient(s). Histologically, 19 cases showed classic findings of PTC, and 2 cases (No. 10 and 14) were follicular variant of PTC (Table 1).
Differentially expressed genes (DEGs) in papillary thyroid cancers
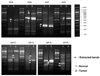
To identify genes that are specifically or predominantly expressed in PTC, we compared the mRNA expression profiles of PTC with those of normal thyroid. To do this, the mRNAs from both samples were extracted and subjected to ACP RT-PCR analysis using a combination of 120 arbitrary primers and 2 anchored oligo(dT) primers (dT-ACP 1 and dT-ACP 2). The expression levels of certain mRNAs were compared between normal thyroid tissue and PTC tissue of same patients. The expression levels of certain mRNAs were either lower or higher in PTCs than in the normal thyroid. The analysis generated about 800 amplicons. There are 8 genes over-expressed in normal thyroid and 9 genes over-expressed in PTC (Fig. 1). Among the 17 DEGs differentially expressed, 10 DEGs isolated were cloned, sequenced, and searched with GeneBank (Fig. 2). These differential display patterns between normal thyroid and PTC, as assessed by ACP RT-PCR, were very reproducible.
Detection of gene from cloning and sequencing
The mRNAs of the DEGs showing distinct results were extracted, amplified using the TOPO TA Cloning kit (Invitrogen, K4500-01, CA, USA) and were sequenced.
The DNA sequence of each gene was confirmed by comparison with sequences in GenBank (NIH, MD, USA). Table 2 shows 10 DEGs accessed by GeneBank and their homologies.
Confirmation of ACP observations by RT-PCR
To confirm the efficacy of the ACP system, the differential expression of DEG was confirmed by RT-PCR using each gene specific primer pair. The DEG3 and DEG14 were subjected to confirmation with RT-PCR, because they were distinctively high in tumor tissue. The primers which we designed are shown in Fig. 3. With RT-PCR, we confirmed ACP observation, as shown in Fig. 4. Additionally, we analyzed 21 PTC samples by RT-PCR for DEG3 and DEG14, and observed up-regulated expression in 17 out of 21 (81%) tumors (Fig. 5).
Overexpressed genes not previously associated with PTC
Genes that we found to be over-expressed in PTCs have previously been associated with other cells, tumor cells, stromal cells, or cell cycle control. Examples include type 1 Collagen, A1 (COL1A1),9,10 which has frequently been shown to be over-expressed, as determined by quantitative RT-PCR. Although COL1A1 expression has been demonstrated in tumor cells and tumor-associated stromal cells in multiple cancers,11 COL1A1 has been reported to be elevated in tumor endothelium, compared with normal endothelium,12 suggesting that they play an important role in angiogenesis and formation of desmoplasia in PTC. Dendritic cell-specific transmembrane protein (DC-STAMP)13-17 is preferentially expressed by DC in vitro and ex vivo. DC-STAMP was originally isolated from a cDNA library of human monocytederived dendritic cells and encodes a protein with putative 7-transmembrane domains.
DISCUSSION
Thyroid cancer is the most common endocrine malignancy, and the biological behavior of PTC varies widely. Although the etiology of PTC is not well defined, a strong association with exposure to ionizing radiation exists in a minority of cases.2 Also, it has been suggested the involvement of specific genes, including tumor suppressor genes, oncogenes, apoptosis-associated genes and other predisposing genes. However, genes, signaling pathways, and other basic mechanisms are currently not well characterized. Another issue is the diagnostic and prognostic biomarkers of PTC. The classical oncogenic alterations commonly seen in thyroid cancer include Ras mutations,18,19 RET/PTC rearrangements,20-22 and BRAF mutation.4 The recently discovered activating mutation in BRAF represents the most common genetic alteration in thyroid cancer. Most of the genetic alterations in thyroid cancer exert their oncogenic actions through the activation of the MAP kinase pathway. When constitutively activated, the MAP kinase pathway leads to tumorigenesis.23,24 The discovery of the BRAF mutation in PTC represents one of the most important recent achievement in thyroid cancer research and is of significant clinical potential. Many attempts have been made to identify markers for thyroid carcinoma that can distinguish between benign and malignant lesions. Most have been proven to be not specific enough, including Galectin-3, BRAF, Met, and Cyclin-D1.3,4,7,25
Techniques designed to identify DEGs in tumors have become critical in modern pathology. Differential display, a screening method described by Liang et al.,26 requires PCR using short arbitrary primers. Although this method is simple, rapid, and requires only small amounts of total RNA, many investigators have experienced significantly high false-positive rates and poor reproducibility of results,27,28 because of non-specific annealing by short arbitrary primers. The use of longer primers (18 - 20 bp) makes it possible to increase the annealing temperature to 60℃ after 1 - 4 initial cycles at 40 - 45℃.29 However, the additional tail sequences of longer primers are involved in nonspecific annealing to the cDNA template during PCR cycles at low annealing temperatures.8 Genefishing technique™ using the ACP system, has recently been developed by Hwang et al.8 to screen for DEG transcripts. It uses primers that anneal specifically to the template and allows only genuine products to be amplified, thus eliminating false-positive results. In the present study, we used the ACP system for the identification of DEGs involved in PTC tumorigenesis, and confirmed the expression patterns of 10 DEGs and quantified their expression by RT-PCR. We found type 1 Collagen A1, DC-STAMP and other novel genes in PTCs.
Collagen type 1 (COL1A1)9,10 has frequently been found to be over-expressed, as determined by quantitative RT-PCR in 17 out of 21 (81%) PTCs. Although COL1A1 expression was demonstrated in tumor cells and tumor-associated stromal cells in multiple cancers,11 COL1A1 was reported to be elevated in tumor endothelium, compared with normal endothelium,12 suggesting that they play an important role in angiogenesis and formation of desmoplasia. Since a dense demoplastic reaction and matrix remodeling induced by tumor are prominent components of most PTCs, over-expression of COL1A1 can be associated with invasive process.
Dendritic cell-specific transmembrane protein (DC-STAMP)13-16 is preferentially expressed by DC in vitro and ex vivo. DC-STAMP was originally isolated from a cDNA library of human monocytederived dendritic cells and encodes a protein with putative seven-transmembrane domains. The pathologic significance of this novel gene in PTC can not currently be explained. Further investigations using immunohistochemistry or in situ hybridization is mandatory for this interesting molecule in PTC.
The expression of calcyclin (DEG1) was studied for in various benign and malignant thyroid lesions by Nagy et al.30 However, its expression in PTC still remains to be evaluated in large series of PTCs.
The other genes differentially expressed in PTCs include SDR member 3 (DEG2),31 PRP8 (DEG4), TIMP1 (DEG7),32 and Cystatin F (DEG13). They have not been studied in PTCs so far, and need further evaluation.
With extensive study for the differential gene expression patterns of PTCs using GeneFishing™ with ACP primers, we conclude as follows;
1) The ACP-based strategy which we used to identify DEGs in PTCs is easy, and produces no false positive and yields reproducible results.
2) With this method, we detected 17 genes that are differentially expressed in PTCs and cloned with sequences of 10 genes.
3) We confirmed the expression patterns of 2 DEGs by RT-PCR assay, and identified consistent results in 81% (17/21) of PTCs.
4) The two DEGs overexpressed in PTCs were confirmed to be DC-STAMP and type I collagen A1. They are novel genes, identified first in PTCs.
5) Although the detailed functions of these two genes and their products remain to be determined, the genes presently identified are expected to provide insights into mechanisms of carcinogenesis or tumor progression in PTCs.






 XML Download
XML Download