

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Cervical cancer is a virus-induced disease that is caused by the integration of a human papilloma virus (HPV) DNA into the host's genome.1-3 Infection with HPV causes disruption of the host's E2 gene, resulting in expression of viral oncogenes, E6 and E7. The E6 and E7 products inhibit the activities of tumor suppressors, p53 and retinoblastoma protein, respectively. This then eventually leads to the accumulation of damaged DNA and the development of cervical cancer.4-5 Although cervical cancer is 1 of the causes of highest mortality in female cancer patients worldwide, it is a curable disease when diagnosed at an early stage.6 However, clear prognostic factors for cervical cancer development are not yet in existence.
Mitochondria, which are cell organelles involved in the processes of cell life and death and in tumoral transformation, appear to have prominent dysfunction in cancer cells. Mitochondrial failure induces abnormal ultrastructures, deregulated metabolism, altered biochemistry and mutation of mitochondrial DNA (mtDNA) in cells.7 Mitochondrial molecular chaperones play important roles in protein transport, protein complex assembly, refolding of misfolded proteins and triggering of protein degradation by proteosomes.8-9 Heat shock proteins (HSPs) are molecular chaperones that are classified into families according to their molecular weight (i.e., HSP100, HSP90, HSP70, HSP60, and small HPSs). HSPs, an evolutionary conserved protein, are ubiquitous and have multiple functions in cellular homeo-stasis including gene expression regulation, DNA replication, signal transduction, differentiation, apoptosis, and cellular senescence or immortalization. They also protect cells from various stresses such as hypoxia or ischemia, as well as sudden increases in temperature.10-12 In normal cells, HSP60 is mostly localized in the mitochondrial matrix and outer mitochondrial membrane, constitutively expressed under normal condition, and induced by heat shock, mitochondrial damage, and mtDNA depletion.13,14 Recently, other molecular roles for mammalian HSP60 have been reported. For example, human HSP60 may trigger apoptosis through caspase cascade activation by an association between HSP60/HSP10 complex and pro-caspase-3 inside the mitochondria, resulting in a subsequent release of the HSP60 into the cytoplasm.15
Overexpression of HSP60 has been reported in various tumors or cancers, such as adrenal Cushing tumors, human breast, large bowel, bronchial, exocervical, ovarian and prostate cancers.16-23 Recently, an upregulation of HSP60 in preinvasive lesions of the cervix has been shown by immuhistochemistry.24 Although HSP60 plays important roles in various biological events, the exact molecular roles of HSP60 are still poorly understood, and the relationship between HSP60 and invasive cervical cancer has not been reported yet. In this study, we investigated the HSP60 expression in invasive cervical cancer tissues and evaluated any prognostic significance of HSP60 in cervical cancer using proteomics, reverse transcriptase polymerase chain reaction (RT-PCR) and Western blot analyses.
MATERIALS AND METHODS
Tissue preparation
The cervical cancer patients were recruited at the outpatient clinic of the Department of obstetrics and gynecology, Gil Medical Center, Incheon, Korea. Tissue samples were collected from 20 cervical cancer patients (mean age, 47.95 ± 16.1 years) and 20 normal controls (mean age, 46.80 ± 12.9 years). Cervical biopsies were obtained from patients with a diagnosis of cervical cancer or uterine myoma as described previously.25 The specimens were brought to clinicopathology department immediately after resection, and parts of the tissues were subsequently dissected, divided into several tubes, placed in liquid nitrogen and stored at -80℃ until analyzed. Cancer samples were characterized according to International Federation of Gynecology and Obstetrics (FIGO) stage as fellows: stage I, 6 case (30%); stage II, 10 case (50%); stage III, 2 case (10%); and stage IV, 2 case (10%). Histopathology of cancer samples was evaluated as fellows: squamous cell carcinoma (SCC), 16 cases (80%); adenocarcinoma, 3 cases (15%); and malignant mixed müllerian tumor (MMMT), 1 case (5%). Patients with cervical cancer underwent radiotherapy and/or chemotherapy after biopsy. Normal cervical tissues came from uterine myoma obtained from women diagnosed benign condition by Pap smear test. Our study was approved by the International Review Board on the experimental studies.
Two-dimensional gel electrophoresis
Biopsy tissues were homogenized in a R/S buffer {9 M Urea, 2% 3-[(3-Cholamidopropyl) dimethylammonio]-1-propanesulfonate, 50 mM dithiothreitol (DTT), 0.4% ampholyte} containing 0.5% each of protease and phosphatase inhibitor cocktail, and then ultracentrifuged at 100,000 g for 1 hour (h) at 20℃. Protein contents of the supernatants were determined using a protein assay kit (BioRad, Hercules, CA, USA), and then the samples were prepared at 1 mg/350 µL concentration with R/S buffer and stored at -80℃. One milligram (mg) of the protein sample was loaded on an immobilized pH gradient (IPG) strip (pH 4-7) in PROTEAN IEF CELL (BioRad, Hercules, CA, USA), and the strip was then covered with mineral oil and rehydrated at 50 V for 12 h. Moist wicks were placed at both ends of the strip to avoid salt contamination while the protein samples were separated by their isoelectric points (IP) at 10 KV for 15 h. The IPG strip was then soaked in equilibration buffer [(375 mM Tris, 6 M Urea, 2% sodium dodecyl sulfate (SDS), 20% glycerol, 2.5% iodoacetamide)] for 10 minutes (min), and then the samples which had previously been separated by IP were separated again on a 2-dimensional SDS-polyacrylamide gel (2D-PAG) [4.0 mL of 30% acrylamide/Bis, 2.5 mL of 1.5 M Tris-HCl (pH 8.8), 0.1 mL of 10% SDS (w/v), 50 µL of 10% ammonium persulfate, 5 µL of N,N,N'-tetramethylethylenediamine in 10 mL of 12% 2D-PAG] by their molecular weights at 20 mA/gel for 16 h. Protein spots on 2D-gel were stained with Coomassie brilliant blue for 24-48 h,26 de-stained in double distilled water (ddH2O), and then the stained protein spot images were analyzed using PDQuest software (BioRad, Hercules, CA, USA). The protein spots that showed an increased staining density of up to two fold compared with their controls were selected for further analysis.
Selected protein spots were individually excised into new tubes, and the Coomassie blue staining was destained with 50% acetonitrile/25 mM ammonium bicarbonate. Gel slices containing protein spots were treated with 10 mM DTT at 56℃ for 30 min, and with 55 mM iodoacetamide at room temperature for 25 min in dark. They were then digested with trypsin at 37℃ for 16 h with shaking. After pooling the supernatant into new tubes, peptide samples were dried completely using SpeedVac system for 4 h. Samples were dissolved in 50% acetonitrile/ 0.1% trifluoroacetic acid (TFA), co-crystallized by mixing with matrix (α-cyano-4-hydroxy-cinnamic acid saturated with 0.1% TFA/50% acetonitrile) and loaded on the silcon-coated 96 well microtiter sample plate (Applied Biosystems, Foster City, USA).
Peptides masses were analyzed using Voyager DE Matrix assisted laser desorption ionization-Time of flight mass spectrometry (MALDI-TOF) mass spectrometer (Applied Biosystems, Foster City, USA). Mass spectra were obtained by averaging 100 to 150 individual laser shots. Calibration of spectra was performed externally by 2 standard peptide, angiotensin 1 (m/z 1296.6853) and adrenocorticoptrophic hormone (18-39 clip) (m/z 2465. 1989). A database search for protein sequence for homology was performed using MS-Fit search algorithm (http://prospector.ucsf.edu/prospector/4.0.8/html/msfit.htm). Mass tolerance for the monoisotopic peptide masses were set to 50 ppm.
Proteomic analysis (i.e., two-dimensional gel electrophoresis and MALDI-TOF analysis) was carried out twice with different 2 samples in each group in order to confirm the peptide profiles analyzed.
Reverse transcription-polymerase chain reaction
Total RNA was extracted from cervical cancer or normal tissues with TRIzol reagent (Invitrogen, Carsbad, CA, USA) according to the manufacturer's instructions. The yield of total RNA was determined by measuring the absorption at 260 nm. Reverse transcription with 1 µg of total RNA was performed in a final volume of 20 µL using 200 U of Superscript II reverse transcriptase, 0.5 µg oligo dT12-18 as a primer, 0.5 mM dNTP Mix, 10 mM DTT, and first strand buffer (Invitrogen, Carsbad, CA, USA). The mixture of oligo dT12-18 primer, total RNA and diethyl pyrocarbonate (DEPC) treated ddH2O was heated at 70℃ for 10 min initially, and other components were then added and incubated at 42℃ for 1 h. Subsequent incubation at 70℃ for 15 min was carried to inactivate the reverse transcriptase.
To determine the relative levels of HSP60 messenger RNA (mRNA), PCR of complementory DNA (cDNA) was carried out in 20 µL mixture containing 1 µL of cDNA, 10X reaction buffer, 2.5 mM decxynucleoside triphosphate (dNTP) mix, 6 pmol each of 5' and 3' primer, and 1U of G-Taq DNA polymerase (Bioneer, Daejeon, Korea); Sense: 5'-AGA TGG AGT GGC TGT GCT GA-3', Antisense: 5'-CAT CAT AAC CAA CTT CTG AG-3'. The reactions were started at 94℃ for 5 min and amplified for 30 cycles of 30 sec at 94℃, 30 sec at 58℃ and 30 sec at 72℃. Final extensions were done for 7 min at 72℃ to compete polymerization. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as an internal control to confirm equal loading of the samples. The PCR products were separated on 2.5% agarose gel using NeSieve GTG agarose (FMC, Rockland, ME, USA), and analyzed by a digital image analysis system.
Western blot
For Western blot analysis, cervical tissues were removed immediately, transferred to LN2 and stored at -80℃. The tissues were homogenized in a single detergent lysis buffer (50 mM Tris, pH 8.0; 150 mM NaCl; 1% Triton X-100; each of 0.5% protease and phosphatase inhibitor cocktail) and then centrifuged at 13,000 g for 20 min at 4℃. The supernatants transferred into new tubes were measured for protein contents using a protein assay kit (Biorad, Hercules, CA, USA), aliquoted at an 5 µg/20 µL concentration in lysis buffer, and stored at -80℃. Otherwise, they were used in the same day. The samples were mixed with loading buffer (100 mM Tris, pH 6.8; 200 mM DTT; 4% SDS; 20% glycerol; 0.2% bromophenol blue) at 1 : 1 dilution, boiled for 5 min, quickly chilled on ice, and then separated on 10% SDS-polyacrylamide Tris-glycine gels. The proteins were then transferred onto nitrocellulose membranes (Hybond-C, Amersham, Bucks, UK) and treated with 5% nonfat dry milk in 1X phosphate buffered saline-Tween (PBST) (1.46 mM NaH2PO4H2O; 8.05 mM Na2HPO4; 144.72 mM NaCl; 5% Tween 20) overnight at 4℃. The membranes were reacted with rabbit anti-HSP60 antibodies (Santa Cruz Biotechnology, CA, USA) at 1 : 1000 dilution for 1 h, and then reacted with HRP-conjugated goat anti-rabbit antibodies (1 : 1000 dilution; Amersham, Bucks, UK) for 1 h at room temperature. The bound antibodies were detected with chemilu-minescence according to the manufacturer's instructions (NEN Life Science, MA, USA), and quantified using a digital image analysis system.
Statistical analysis
A one-way analysis of variance (ANOVA) was used to analyze the expression of HSP60 in cervical cancer and normal tissues. RT-PCR and Western blot data were subjected to post-hoc Fisher's protected least significant difference (PLSD) test and presented by means ± SEM. For all comparisons, the level of significance was set at p ≤ 0.05.
RESULTS
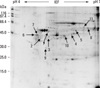
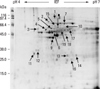
We performed 2D proteomic analysis to evaluate the differences in protein expression between human cervical cancers and normal cervical tissues. Protein spots on the 2D gels were stained with Coomassie blue, and the staining densities of the spots were analyzed with PDQuest software. Proteomic analysis was carried out twice with different 2 samples from each group in order to confirm the peptide profiles analyzed and one of them is presented (Figs. 1 and 2). In 2D gel analysis of normal cervical tissue, 11 spots with higher staining densities then cancer tissue were selected for MALDI-TOF assay (Fig. 1) and identified (Table 1). Fig. 2 reveals the 2D gel image of the cervical cancer tissue. Nineteen spots of cervical cancer tissue samples that showed up to 2 fold increased staining density compared with their normal control were selected and identified (Table 2). Specifically, HSP60 protein (Spot 6) which was overexpressed on cervical cancer gel displayed dramatically high 98.2% of sequence and corresponded to its intact molecular size (60 kDa) (Table 2).
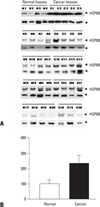
In order to ascertain the increase of HSP60 expression in cervical cancer tissues, we investigated HSP60 mRNA expression in cervical cancer and normal cervical tissues obtained from twenty cancer patients and twenty normal controls, using semi-quantitative RT-PCR (Fig. 3A). All samples were quantified and corrected for total input RNA by normalizing the expression value of GAPDH. Although HSP60 mRNA was detected in both groups, no statistically significant differences between cervical cancer and normal cervical tissues were noted (Fig. 3B).
To examine the expression levels of HSP60 (60 kDa) protein, we carried out Western blot analysis in both groups (Fig. 4A). Anti-actin (43 kDa) was used as an internal control and expression of HSP60 was normalized by actin protein. The results obtained from twenty cases of each group showed that the expression of HSP60 protein in cervical cancer tissues was significantly increased compared to cervical normal tissues (p < 0.05) (Fig. 4B).
DISCUSSION
The immunohostochemical evaluation of HSP60 in precancer of the cervix has recently been reported,24 however the relationship between HSP60 and invasive cervical cancer has not yet been reported. In this study, we compared expression levels of HSP60 mRNA or protein between invasive cervical cancers and cervical normal tissues.
2D gel proteomic analysis revealed that the expression of HSP60 was increased in cervical cancer tissues compared to normal tissues (Figs. 1 and 2). It is generally accepted that DNA damages which cause cancer induce cell cycle arrest, apoptosis or DNA mutation, resulting from changes in protein expression. In the present study, protein spots on the 2D gels showed different expression patterns between cervical cancer and normal cervical tissues, indicating that the protein expression of normal cervical tissues was changed by induction or inhibition of genes, and/or by degeneration or modification of proteins during carcinogenesis. HSP60 protein of cervical cancer tissues was markedly elevated, revealed by 2D gel proteomics and validated by Western blot (Fig. 4). However, there was no difference in HSP60 mRNA levels between cancer and normal tissues (Fig. 3). These findings suggest that serial processes after transcription become important step in cervical cancer development. Indeed, differences of HSP60 expression were detected in both 2D gels and Western blot, indicating difference in protein level, but not in mRNA level.
It has been known that HSP60 is constitutively expressed under normal conditions, and its expression is induced by stressful conditions such as heat shock, mitochondrial damage, and mtDNA depletion.14 Although the expression of HSP60, which is localized in mitochondria, is directly related to mitochondrial regeneration after cell division or the increase of mitochondrial activity during normal conditions, and HSP60 is significantly less expressed in normal tissues.13,18,27 On the other hand, the present study found that HSP60 was significantly overexpressed in cervical cancer tissues compared to normal cervical tissues. The overexpression of HSP60 in cervical cancer tissues suggests that this protein might play a different role in cervical carcinogenesis. Cappello et al.22 reported that elevated expression of HSP60 may be a protective upregulation against cancer development (i.e. the blockade of apoptotic machinery that usually takes place during cancer progression). The involvement of HSP60 in the process of apoptosis and tumorigenesis is still in dispute. An antiapoptotic effect of HSP60 and down-regulation of HSP60 have been reported in cardiac myocytes and bronchial cancer, respectively.28,29 On the other hand, overexpression of HSP60 has been reported in breast and ovarian carcinomas and myeloid leukemia.30-32 Moreover, recent studies showed upregulation of HSP60 during carcinogenesis of the large bowel and the uterine exocervix.18,22 Specifically, overexpression of HSP60 has been demonstrated in early cervical carcinogenesis as well as in prostate and squamous cervical cancers of both early and advanced grades.18,24,27 These results support the possibility that the expression of HSP60 may play a role as a prognostic factor of cancer development. By employing immunohistochemistry, Castle et al.24 showed up-regulation of HSPs in response to HPV infection and early cervical carcinogenesis, and discussed the role of HSP60 and HSP70 as surrogate marker for precancer development. However, the present study found the presence of HPV type 16 in 75% of our cervical cancer samples, demonstrated by PCR (data not shown) and they were shown to be invasive cancer stage I to IV, suggesting that HSP60 expression is also upregulated in invasive cervical cancer stages as well as precancer stage.
In conclusion, we compared HSP60 mRNA level and protein expression in cervical tissues diagnosed as normal and invasive cervical cancer, and confirmed upregulation of HSP60 protein in invasive cervical cancer tissues. It suggests that HSP60 is involved in cervical carcinogenesis, and can be used as a useful prognostic tool. Since the role of HSP60 in cancer progression and mechanisms involved in the regulation of HSP60 expression remain unclear, more studies are needed to elucidate the relationship between HSP60 and cancer development.


 XML Download
XML Download