

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
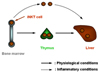
Facultative intracellular bacteria comprise various pathogens such as Mycobacterium tuberculosis, M. leprae, Salmonella enterica, Brucella sp., Legionella sp., Listeria monocytogenes, and Francisella tularensis, all of which can survive in professional phagocytes such as macrophages.1 Although innate immunity orchestrated by multiple cell populations, including granulocytes and macrophages, is pivotal for the elimination of these bacteria, conventional T cells are mandatory for sterile eradication of these pathogens (Fig. 1).1 Because the majority of facultative intracellular bacteria are trapped in the liver immediately after systemic infection, T cells that reside in and/or infiltrate the liver should play a decisive role in the following course of infection (Fig. 1). Experimental murine listeriosis models are instrumental in analyzing the role of T cells in the liver during facultative intracellular bacterial infection, not only because protection against L. monocytogenes strictly depends on T cells, but also because liver parenchymal cells serve as a reservoir for this bacterium (Fig. 1).1-3
The liver is a rich provenance of unconventional T cells, called natural killer (NK)T cells, co-expressing NKR-P1B/C (NK1.1)(CD161) that are type II membrane glycoproteins of the C-type lectin superfamily.4 The majority of NKT cells express an invariant (i) T cell receptor (TCR), typically comprising Vα14/Jα18 combined with a highly skewed TCRVβ towards Vβ8.2 in mouse, and homologous chain Vα24/Jα18 paired with Vβ11 in human (iNKT cells).4 The liver iNKT cells have a great potential to secrete both type 1 and type 2 cytokines.4-7 The high abundance of iNKT cells in the liver and their rapid and vigorous cytokine release in response to stimuli suggest participation of this cell population as an immunomodulator in the liver.
iNKT cells have been shown to participate in the regulation of various immune responses; e.g. tumor rejection8,9 and prevention of the development of autoimmune diseases.10-12 Although iNKT cells have been suggested to participate in elimination of various microbial pathogens,13-26 recent studies argue against the crucial role of this cell population in some microbial infections.27-32 Moreover, new studies have shed light on the intriguing aspects of the NKR-P1 family, including NKR-P1B/C (NK1.1), in controlling immune responses.33-40 Thus, iNKT cells appear to play more complicated roles than originally thought. Here, we focus on the unique aspects of iNKT cells as regulatory cells during murine listeriosis and the role of NK1.1 expressed on these cells.
Is NK1.1 a reliable marker for iNKT cells?
Although iNKT cells were originally regarded as T cells co-expressing NK1.1, this cell population does not seem to consistently express this marker.41-44 Immature iNKT cells lack surface expression of NK1.1, but they acquire the marker expression during ontogeny, suggesting that the NK1.1- subset is a precursor of NK1.1+ subpopulation.43,44 Yet, substantial numbers of iNKT cells lacking NK1.1 have been identified in the periphery.28,41,42 This suggests that NK1.1 is not merely a marker for mature iNKT cells and raises the possibility that NK1.1 surface expression on iNKT cells is fluctuated under various conditions.
iNKT cells become undetectable upon activation.6,7,42,45-61 Although the disappearance of iNKT cells had been considered to be caused by activation-induced cell death/apoptosis (Fig. 2A),48,51,53,61 recent studies suggest that iNKT cells robustly expand in situ rather than undergoing apoptosis.57-59: i.e. the failure of iNKT cell detection is caused by the loss of NK1.1 and TCR, which were previously considered reliable markers for the detection of iNKT cells (Fig. 2A).57-59 Yet, the loss of surface expression of NK1.1 and TCR, and subsequent re-expression of marker(s) have thus far been observed only in iNKT cells stimulated with their agonist, α-galactoceramide (α-GalCer).57-59
Fluctuation of liver iNKT cells during L. monocytogenes infection
Cells stained with monoclonal antibodies (mAbs) against surface markers, including NK1.1 and TCR, become transiently undetectable in the liver of mice following L. monocytogenes infection.6,46 This dynamic fluctuation of iNKT cells during L. monocytogenes infection has recently been verified using α-GalCer-loaded CD1d tetramers (α-GalCer/CD1d tetramers): the numbers of α-GalCer/CD1d tetramer-reactive T cells co-expressing NK1.1 (NK1.1+ iNKT cells) are markedly reduced in the liver during the early stages of L. monocytogenes infection, whereas those of α-GalCer/CD1d tetramer-reactive T cells lacking NK1.1 (NK 1.1- iNKT cells) rise following infection and become dominant among the iNKT cell population (Fig. 2B).28 Similar kinetics of iNKT cells are seen in Vβa mice, which are devoid of iNKT cells expressing TCRVβ8,28 implying that the curtailment of the NK1.1+ subset and subsequent expansion of NK 1.1- subpopulation occur independently from TCRVβ usage. Hence, iNKT cells are markedly influenced by L. monocytogenes infection, and the NK1.1+ and NK1.1- subsets of iNKT cells show differential kinetics during listeriosis.
Participation of endogenous interleukin-12 in fluctuation of liver iNKT cells during L. monocytogenes infection
Interleukin (IL)-12 secreted mainly from macrophages, dendritic cells, and granulocytes is a heterodimeric cytokine which is composed of a covalently linked 35-kDa light chain (p35) and a 40-kDa heavy chain (p40).62 IL-12 stimulates type 1 immune effector functions (e.g. promotion of IFNγ secretion from Th1 cells and NK cells),62 and hence, are mandatory for protection against intracellular microorganisms including L. monocytogenes.2,63
Neutralization of endogenous IL-12 (p40) reverses the curtailment of the NK1.1+ subset and the subsequent expansion of the NK1.1- subpopulation of iNKT cells during listeriosis.28,46 The kinetics of NK1.1+ and NK1.1- iNKT cells in the liver following L. monocytogenes infection is paralleled by numerical changes of IL-12 producers in the liver.28,64,65 It is thus conceivable that fluctuation of iNKT cells during listeriosis is determined by IL-12 (p40) levels in the hepatic microenvironment.
Although IL-12 (p40) had been considered a useful tool for determination of the role of IL-12 in vivo, the p40 subunit of IL-12 has been demonstrated to be shared by IL-23, a cytokine which is a disulfide-bridged complex of a p19 subunit and the p40 subunit of IL-12.62,66 It is therefore possible that IL-23 rather than IL-12 participates in the fluctuation of iNKT cells during L. monocytogenes infection. Yet, similarly to L. monocytogenes infection, numbers of NK1.1+ subset are markedly diminished following in vivo administration of recombinant (r)IL-12, and the NK1.1- subpopulation expands thereafter (Fig. 2B).28 Thus, it is tempting to assume that endogenous IL-12 plays a central role in the fluctuation of iNKT cells during listeriosis, although it cannot completely be excluded that IL-23 also, at least in part, participates in this mechanism.
Fluctuation of iNKT cells is differentially controlled by signaling through TCR and IL-12R
iNKT cells become undetectable after stimulation with their agonist, α-GalCer.42,49,50,54,56-59 In contrast to L. monocytogenes infection, this change is not prevented by IL-12 neutralization (Fig. 2A).64 Thus, endogenous IL-12 is involved in the disappearance of NK1.1+ iNKT cells during L. monocytogenes infection (Fig. 2B),28,46 whereas the disappearance of NK1.1+ iNKT cells by α-GalCer treatment apparently occurs independently from IL-12 (Fig. 2A).64
The TCR surface expression on iNKT cells is down-modulated after α-GalCer stimulation (Fig. 2A),57-59 whereas that on iNKT cells is not affected by Salmonella infection.58 Similar to L. monocytogenes infection, fluctuation of iNKT cells during salmonellosis is also controlled by endogenous IL-12 [Emoto, unpublished observation]. It is therefore assumed that disappearance of NK1.1+ subset and subsequent expansion of NK1.1- subpopulation are differentially controlled by signalling though TCR and IL-12R.
Origin of NK1.1- iNKT cells which emerge in the livers of mice following L. monocytogenes infection
The numerical increase of liver NK1.1- iNKT cells after L. monocytogenes infection is prevented by NK1.1+ cell depletion.28 This suggests that NK1.1+ cells are a prerequisite for the emergence of NK1.1- iNKT cells. Because NK1.1+ cells comprise not only iNKT cells, but also NK cells, it is possible that the numerical increase of NK1.1- iNKT cells following L. monocytogenes infection is controlled by NK cells. However, because numerical increase of NK1.1- iNKT cells following L. monocytogenes infection is further increased by NK cell depletion (anti-asialo GM1 Ab treatment),28 this possibility is unlikely.
In addition to NK and iNKT cells, NK1.1+ cells comprise nonclassical NKT cells. Although iNKT cells comprise cells expressing CD4 but lacking CD8 (CD4+ cells), and those lacking CD4 and CD8 (CD4-8- (double negative: DN) cells), a small but distinct NKT cell population expressing CD8 has been identified.67-71 Moreover, in contrast to iNKT cells which express TCRα/β as an antigen receptor, some NKT cells expressing TCRγ/δ have also been identified.70,72-75 Because both cell populations are also abundant in the liver70,75 and are depleted by NK1.1+ cell depletion, it is possible that the prevention of the emergence of NK1.1- iNKT cells by NK1.1+ cell depletion is caused by the depletion of these cells. However, because numerical increase of liver NK1.1- iNKT cells following L. monocytogenes infection is not prevented by CD8α+ or TCRγ/δ+ cell depletion,28 this possibility is also unlikely. Thus, the NK1.1- subset, which emerges in the liver of L. monocytogenes-infected mice, is primarily derived from the NK1.1+ subpopulation of iNKT cells.
Do iNKT cells propagate in situ following L. monocytogenes infection?
Recent studies suggest that iNKT cells expand in situ robustly in response to α-GalCer.57-59 It is possible that a similar mechanism exists in listerial infection, because total numbers of iNKT cells are increased, though transiently reduced, following L. monocytogenes infection compared to preinfection.28 However, because (i) numbers of NK cells are increased in the liver following L. monocytogenes infection,27,28 (ii) expansion of iNKT cells is impaired in the presence of NK cells,76,77 and (iii) expansion of liver iNKT cells is promoted by NK cell depletion, which is caused by increased local concentrations of IL-1528,76 (IL-15 is prerequisite for the proliferation of not only NK cells but also iNKT cells78), it is conceivable that iNKT cells increased in the liver following L. monocytogenes infection are supplied from other organs. Hence, it is likely that stimulation by a specific antigen (i.e. α-GalCer) and by the cytokine IL-12 have differential outcomes.
Infiltration of iNKT cells in the liver following L. monocytogenes infection occurs independently from thymus
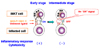
Although thymus is essential for the development of iNKT cells (Fig. 3),5,70,79-81 comparable numbers of those cells are detected in adult thymectomized mice.28,82 Similar to euthymic mice, numbers of the NK1.1+ subset are markedly diminished in adult thymectomized mice following L. monocytogenes infection, and the NK1.1- subpopulation is numerically increased.28 Thus, numerical alterations in NK1.1+ and NK1.1- iNKT cell populations in response to L. monocytogenes infection occur independently from a functional thymus. The NK1.1+ subset in the periphery has been found to be derived from a thymic NK1.1- subpopulation of iNKT cells.43,44 In this study, NK1.1 surface expression on iNKT cells is acquired in the periphery after the cells have left the thymus during ontogeny. However, the NK1.1- subset that emerges in the liver following L. monocytogenes infection develops from the NK1.1+ subpopulation of iNKT cells.28 Because the disappearance of liver iNKT cells after TCR stimulation has been shown to be followed by repopulation of these cells due to homeostatic proliferation of an iNKT-cell reservoir in the bone marrow,48 it is conceivable that some NK1.1+ iNKT cells, which re-emerge at later stages of listeriosis, are derived from the bone marrow, although in situ expansion of iNKT cells cannot completely be excluded (Fig. 3). Hence, accumulation of iNKT cells in the liver is differentially regulated under physiological and inflammatory conditions.48
Functional alterations of liver iNKT cells during listeriosis
Substantial numbers of IFN-γ and IL-4 producers are detected among liver iNKT cells from uninfected mice after in vitro stimulation with TCR/CD3 ligation5,6 or with phorbol myristate acetate and ionomycine28 (Fig. 4). High numbers of IFN-γ producers are detected among liver iNKT cells from L. monocytogenes-infected mice, whereas IL-4-producing cells are virtually undetectable (Fig. 4).6,28,46 Because the vast majority of iNKT cells in the liver of L. monocytogenes-infected mice lack surface expression of NK1.1, these findings suggest that NK1.1- iNKT cells, which emerge in the L. monocytogenes-infected liver, fail to produce IL-4 and hence express a Th1-like phenotype.
In mouse, the iNKT cells segregate into 2 populations on the basis of CD4 expression; i.e. CD4+ and DN cells.4,5,83 Before infection, the majority of liver iNKT cells co-express CD4 and only a minority lack this marker.5,28 The CD4+ iNKT cells are numerically reduced after L. monocytogenes infection, whereas DN iNKT cells are virtually unaffected (Fig. 4).6,28,46 Hence, L. monocytogenes infection primarily compresses the CD4+ rather than DN iNKT-cell population.
Since the CD4+ and DN iNKT cells are differentially influenced by L. monocytogenes infection, it is possible that each cell subset plays a different role during L. monocytogenes infection. In uninfected mice, frequencies of IL-4-producing iNKT cells are markedly reduced by CD4+ or NK1.1+ cell depletion, whereas numbers of IFN-γ producers are virtually unaffected.5,28 In contrast, frequencies of IFN-γ-producing iNKT cells are markedly higher in CD4+ cell-depleted mice after infection as compared to nondepleted mice.28 These findings suggest that NK1.1- iNKT cells with IFN-γ-producing activity in the L. monocytogenes-infected liver are preferentially DN (Fig. 4).
DN iNKT cells differ from CD4+ iNKT cells in their cytokine production profile.69,84-88 In general, CD4+ rather than DN iNKT cells are responsible for IL-4 production albeit at varying levels in different organs. Since (i) CD4+ cells dominate DN cells among liver iNKT cells,4,5,28 (ii) IL-4-producing cells in the liver are markedly reduced by NK1.1+ cell depletion,6,46 (iii) large numbers of IL-4 producers are detected among purified liver CD4+ NK1.1+ cells after TCR ligation,6 and (iv) numbers of IL-4-producing iNKT cells from CD4+ or NK1.1+ cell-depleted mice are minute,6,28 it appears that the CD4+NK1.1+ subset under physiological conditions is mainly responsible for IL-4 production in the liver although iNKT cells express both IL-4 and IFN-γ mRNA upon stimulation.89,90 Differential stimulation (i.e. specific antigen versus IL-12) is probably responsible for distinct cytokine production with inflammation, thus driving IFN-γ production from iNKT cells.5,28,46
NK1.1- iNKT cells re-express NK1.1 at later stages of listeriosis
Numbers of iNKT cells reach levels comparable to those in uninfected animals at later stages of listeriosis.46,64 In parallel, the NK1.1+ subset is proportionally increased and NK1.1- subpopulation is reduced. It is thus possible that NK1.1 is re-expressed on iNKT cells even if the marker was lost. Substantial numbers of donor-derived NK1.1+ iNKT cells are detected in the liver of recipient re-arrangement gene-1-/- mice lacking all T cells, including iNKT cells, which was reconstituted with NK1.1+ cell-depleted hepatic leukocytes from mice that had been infected with L. monocytogenes.28 These findings suggest that NK1.1 can be re-expressed on iNKT cells, even in spite of previous loss of the marker (Fig. 2B), although it cannot completely be excluded that NK1.1- iNKT cells fail to accumulate in the liver of recipient mice. It is conceivable that at least NK1.1+ iNKT cells, which re-emerge during late stages of listeriosis, express functional activities similar to those in naive mice, because NK1.1+ iNKT cells that re-emerged in the liver have a potential to secrete IL-4.64
Re-expression of NK1.1 on liver iNKT cells from mice, which had been infected with L. monocytogenes, is not found after in-vitro culture even in the presence of IL-12-neutralizing mAb [Emoto et al., unpublished observation]. This is consistent with previous findings showing that TCR, but not NK1.1, becomes detectable on α-GalCer-stimulated iNKT cells after in-vitro culture.58 These findings suggest that different mechanisms exist in the disappearance/re-emergence of NK1.1 and TCR, and that some factor(s) other than IL-12 also participate in down-modulation of NK1.1 on iNKT cells.
Is down-modulation of NK1.1 on iNKT cells essential prerequisite for combat L. monocytogenes?
The NKR-P1 family comprises activatory and inhibitory receptors. Whereas NKR-P1A, NKR-P1C, and NKR-P1F are activatory receptors, NKR-P1B and NKR-P1D are inhibitory receptors.33-40,91 Lectin-like transcript 1 or C-type lectin-related molecules have been identified as ligands for NKR-P1A, NKR-P1B, NKR-P1D, and NKR-P1F.38-40 Thus, NKR-P1 family members allow recognition of "missing-self", thus controlling activation/inhibition of NK1.1+ cells in a MHC class I-independent manner. Because cross-linking of NKR-P1C by anti-NK1.1 mAb induces IFN-γ production from NK1.1+ cells,33 it is possible that NKR-P1C participates in immunosurveillance such as the elimination of cells lacking hither-to-unknown antigen(s) expressed on infected cells (Fig. 5). It is therefore possible that NK1.1 participates in surveillance of infection, and that the loss of NK1.1 counteracts excessive inflammatory responses.
Detrimental role of iNKT cells against L. monocytogenes infection
IFN-γ plays an essential role in resistance against L. monocytogenes infection,2,92-95 whereas IL-4 exacerbates disease.96-99 Since considerable numbers of IFN-γ, but not IL-4-secreting iNKT cells, are found in the liver of L. monocytogenes-infected mice, it is possible that iNKT cells participate in protection against L. monocytogenes infection. Yet, Jα18-/- mice, which are entirely devoid of iNKT cells, are more resistant to L. monocytogenes infection than control mice.28 These findings suggest that iNKT cells do not participate in antilisterial resistance and may even exacerbate disease, although contribution of iNKT cells in protection against enteric listeriosis has been suggested.100 Because iNKT cells comprise a heterogeneous population, it is speculated that the IFN-γ-producing NK1.1- subset of iNKT cells ameliorates, whereas the IL4-producing NK1.1+ subpopulation exacerbates disease (see Fig. 4).
At first sight, the finding that listeriosis in mice lacking total iNKT cells is ameliorated could be taken as an argument against a pivotal role of the iNKT cells in protection against L. monocytogenes infection. However, 2 subsets of iNKT cells exist: (i) The CD4+NK1.1+ subset which produces IL-4 and hence should be of detriment in listeriosis. At early stages of infection, exacerbation by this subset seems to dominate because depletion of the total iNKT cell population ameliorates listeriosis. This notion is consistent with previous findings showing that listeriosis is improved by anti-CD1 mAb treatment.29 (ii) The CD4-NK1.1- subset produces IFN-γ suggesting its beneficial role in listeriosis. Yet, contribution of the NK1.1- subset to resistance occurs later and seems supportive but not essential.
CONCLUSION
NK1.1 surface expression and functional activities of iNKT cells are markedly influenced by listerial infection. Despite the designation of NKT cells, the NK1.1 surface molecule is not a reliable marker of this cell population. Although iNKT cells produce both IFN-γ and IL-4 in naive mice, the majority of this cell population produces IFN-γ during listeriosis, but not IL-4 due to abundant IL-12 in microenvironment. It is therefore tempting to assume that distinct iNKT-cell populations play different roles in intracellular bacterial infection. Of these, the NK1.1+ subset seems ineffectual or even harmful, whereas the NK1.1- subset appears to contribute to antilisterial protection by means of IFN-γ. NK1.1 surface expression on iNKT cells in the liver fluctuates during L. monocytogenes infection in a reversible manner. This dynamic fluctuation of NK1.1 expression on iNKT cells suggests a unique role of the NK1.1 molecule on this cell population during intracellular bacterial infection.



 XML Download
XML Download