

 PDF
PDF ePub
ePub Citation
Citation Print
Print


Over the last 20 years, new and reemerging pathogens (e.g., West Nile Virus, anthrax) have appeared at alarming rates, garnering considerable attention from the scientific and public health communities, as well as from the media. The Chinese, who recently dealt with the terror caused by Severe Acute Respiratory Syndrome (SARS), now face the threat posed by avian influenza. Avian influenza is commonly known as "bird flu" and has caused enormous economic loss for China.1 This review will discuss the features of highly pathogenic avian influenza viruses (H5N1), incidence, infection, immunity, clinical management, prevention, and control in Mainland China.
THE FEATURES OF H5N1
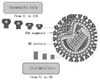
Avian influenza virus is a single-stranded RNA virus of the Orthomyxoviridae family. Influenza viruses are classified into groups A, B and C according to differences in the nucleocapsid and matrix protein.2 The type A viruses were divided into haemagglutinin (H) and neuraminidase (N) subtypes based on the surface glycoproteins.3,4 Up until to now, 16 H and 9 N subtypes have been reported (Fig. 1).5,6 All influenza subtypes can be found in waterfowl, but only the H 1-3 and N 1-2 subtypes were known to infect humans; in particular, the H5N1 subtype is highly pathogenic and endemic in Asian nations including Mainland China. In 1997, the H5N1 virus was first isolated from a three-year-old boy in Hong Kong,7 and new genotypes of H5N1 virus have continually emerged in Mainland China.8 H5N1 virus infection has three features. First, the area where avian influenza cases occurred did not have animal plague and the cases of H5N1 showed dissemination in Mainland China.9 Secondly, the outbreak was mainly distributed in Central, East China and South China; in South China the outbreak was more serious. Third, the virus pathogenicity was strong. The H5N1 subtypes not only caused the death of fowl, ducks, goose and turkey, but also caused human morbidity(Fig. 2).10-12
INCIDENCE AND TRANSMISSION
The confirmed cases of avian influenza A (H5N1) in humans since 2003 have been listed in Table 1.13-15 From this table, it was found that the avian influenza plague had been reported in several cities of Mainland China in 2003 and 2004. However, no cases had been diagnosed in humans.16 At the same time it was also found that most cases appeared in females, which may be associated with genotype, although to determine the true reason required further research, and the frequencies of H5N1 infection among humans have not been reported. Human influenza is transmitted by way of the inhalation of infectious droplets and droplet nuclei by direct or indirect contact.17 There were three primary routes for the transmission of avian influenza, the first of which was by contact with the domestic fowl market; the second by bird migrations; and the third was by the transmission from fowl to mammals. Additionally, the infected birds, which were captured, led to poultry infection, although this type of transmission was merely a possibility. Avian influenza usually involves horizontal transmission, while the vertical transmission of the virus has not been discovered. However, the H5N2 subtype has been isolated from chicken eggs, indicating a possibility for vertical transmission.18 For human influenza A (H5N1), the main transmission routes were from birds to humans and from the environment to humans. Human-to-human transmission of influenza A (H5N1) has been suggested in several household clusters, and there was also one case of possible child-to-mother transmission. Intimate contact without the use of protective measures was implicated, and no case of human-to-human transmission by small-particle aerosols was identified.19 In 1997, human-to-human transmission apparently did not occur through social contact, and serologic studies of exposed health care workers showed that the transmission was inefficient. Recently, some researchers have suggested that the animal virus strains may be adapting to humans by accumulating nonsynonymous (amino acid-changing) substitutions in key proteins (e.g. surface glycoprotein).20 However, epidemiologic and virologic studies are needed to confirm whether avian influenza viruses have acquired the capacity for human infection by adaptive evolution. To date, the risk of nosocomial transmission to health care workers has been low.
INNATE IMMUNE RESPONSE TO H5N1
The early stage of an infection is often viewed as a race between the virus and the host's immune system. The first layer of immunity against virus invasion is the integrity of the body surface. Once breached, early 'non-specific' or innate immunity defenses, such as interferons, macrophages and natural killer (NK) cells, become active.21 Type 1 interferons have a primary protective function in the early stages of influenza virus infection.22 Interferons can induce the expression of 'Mx' genes, which can inhibit primary transcription of the viral PB2 polymerase.23,24 To combat this immune response, viruses employ an 'NS' gene that encodes a type 1 interferon antagonist that promotes virus growth.25 Although interferon-γ (IFN-γ) has an obvious function against other viruses, there are no findings to date to support that this cytokine can control influenza viruses.26 Similarly, no involvement of IFN-γ-producing natural killer cell has been identified. However, some evidence indicates that α-galactosylceramide-stimulated natural killer T cells may be of some benefit.27
Evidence showed that the H5N1 virus mainly attaches to type II pneumocytes, alveolar macro-phages and nonciliated bronchiolar cells in the human lower respiratory tract (LRT).28 Some researchers have also found that the number of alveolar macrophages increases during an early influx of neutrophils into the infected lung, which may imply that alveolar macrophages may be protective during the initial stage of infection.29
ADAPTIVE IMMUNE RESPONSE TO H5N1
An absence of T cells renders the host highly susceptible to viral attack. Although our knowledge of avian influenza cellular immunology has expanded rapidly in the last decade, very little was known about the importance of cellular immunity against the avian influenza virus.30 The influenza antigens may travel to the regional cervical and mediastinal lymph nodes or to the spleen by dendritic cells (DCs), which exit at the respiratory tract.31 The amount of antigen-specific response that is generated by direct exposure to migrated DCs or by cross-presentation on resident lymphoid tissue DCs is unclear.32 Studies have found that only the CD8α+ DCs can stimulate influenza peptide-specific T cells or hybridoma cell lines.33 Analysis of the CD8+ T cell response has indicated that all antigen-presenting cells (APCs) could eliminate virus infection within 10-14 days.34 However, experiments using T cell receptor (TCR)-transgenic CD4+ T cells have suggested that APCs are capable of stimulating influenza-specific CD4+ T cells with lower efficiency, and this process can still be occurring for three weeks after initial viral exposure.35,36 No evidence indicates that either viral RNA or antigen can induce immune responses for longer periods.
The humoral immune response in naturally infected poultry includes systemic and mucosal antibody production.37 Antibodies provide a major barrier to virus spread between cells and tissues, especially in restricting virus spread in the blood stream. Antibodies may be generated against any viral protein in the infected cell. Antibodies against glycoproteins expressed on the virion envelop or on the infected cell membrane are of importance in controlling infection.21,38 Influenza viruses produce ten viral proteins that can be divided into three main categories: surface proteins, internal proteins, and nonstructural proteins.39,40 The surface proteins, which are the only antigens capable of inducing neutralizing antibodies, include three types of proteins: Haemagglutinin (HA), Neuraminidase (NA), and Matrix 2 (M2) proteins.41,42
The HA protein has two main functions: one is that of the virus receptor binding site; and the other is used to fuse the viral RNA that is released into the host cell.43 Regarding HA, at least five antigenic epitopes have been discovered for human influenza viruses, and each epitope was capable of inducing neutralizing antibodies. Antibodies against the HA protein were the main determinant for protection in the host against disease, and vaccine for influenza in poultry is primarily based on the HA subtype. The importance of the HA protein requires further investigation.44 The NA protein is an enzymatically active protein that is important in allowing the virus to be released from the cell surface.45 The NA protein also induces neutralizing antibodies in chickens. Antibodies to the NA protein are of less importance than those to the HA protein.46
The M2 protein (M2e) is an integral membrane protein that functions as an ion channel for the virus particle. The antibody to the M2e protein in mice does not provide complete protection, although it does reduce the amount of virus that is shed and provides some protection from the disease. However, the antibody to the M2e protein is well conserved for all influenza type A viruses and potentially could provide protection for all HA and NA subtypes.47 Recent studies found that human influenza vaccines based on the extracellular domain of influenza M2e induced broad-spectrum protective immunity in various antigen constructs.48 The mucosal immune response also probably has a role in protection from H5N1 infection since the initial exposure to the virus is through a mucosal surface. However, little direct work has been done regarding the mucosal immune response in chickens and turkeys.49 Although both antibodies and T cells contribute to the response against influenza virus infections, the viral antigens are less recognizable by the host's acquired immunity because of "antigenic drift".50 As a result of antigenic drift, avian influenza viruses can escape host immune surveillance. An even more radical method of immune evasion occurs when the segmented structure of influenza A genome allows swapping of major gene segments when two different influenza A virus strains infect the same cell.51 At the same time, influenza A virus NS1 protein can not only inhibit innate immunity by preventing type 1 IFN release, but also can inhibit adaptive immunity by attenuating human DC maturation and decreasing the number of DCs available to induce T-cell responses.52,53
CLINICAL MANAGEMENT AND PREVENTION
The current danger to people from avian influenza has been recognized. The World Health Organization (WHO) and Centers for Disease Control and Prevention (CDC) have adopted measures to prevent the emergence of avian influenza in Asia and control its wider transmission.54 The CDC's response has been focused on enhancing surveillance and laboratory testing for human avian influenza (Table 2).55
The Chinese government also exerts great importance on the prevention and control of avian influenza. The Chinese Center for Disease Control and Prevention has published interim guidelines to limit the possibility of human infections during outbreaks of avian influenza in domestic birds and poultry in Mainland China.56-59 In addition, the Chinese government has taken a series of concrete measures in this regard:
THERAPY
Influenza A (H5N1) virus is susceptible to Oseltamivir (Tamiflu) and Zanamivir (Relenza)70 but is resistant to Amantadine and Rimantadine.71,72 Treatment should be started within 48 hours of onset of fever, without waiting for laboratory confirmation.73,74 Mild cases are treated. A higher dose of 150mg twice daily and treatment for 7 to 10 days is required for the treatment of severe infections. Salicylate administration should be avoided in children younger than 18 years to prevent the possibility of Reye's syndrome. The effectiveness of Zanamivir in reducing the severity and duration of illness and in preventing complications has been proven in children from 5 to 12 years old.75
CONCLUSION
The Chinese government learned a lesson during the SARS outbreak. During the threat of avian influenza pandemic, the Chinese Government has taken a series of open, preventive and surveillance measures to control avian influenza transmission.76 Infected fowl have been the primary source of H5N1 infections in humans in Mainland China. At present, transmission between humans is limited, although long-term monitoring is required to identify viral adaptation to human hosts. With increasing poultry production worldwide, it is not a question whether a new pandemic strain will emerge but rather when it will emerge. This matter requires the concerted efforts of poultry producers to improve biosecurity. Governments also will need to intensify their efforts to educate their citizens concerning the risks of avian influenza.




 XML Download
XML Download