

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
While a sympathectomy or a sympathetic blockade relieves neuropathic pain behaviors in sympathetically maintained pain (SMP) conditions, neuropathic pain with no benefit from sympathetic intervention is called sympathetically independent pain (SIP).1 SIP is not only a difficult syndrome to treat, but it is poorly understood.
The L5 and L6 spinal nerve ligation (SNL) model has been reported SMP-dominant neuropathic pain. Neuropathic pain behaviors in the SNL model were relieved by chemical sympathetic block and by surgical sympathectomy.2 Rats with a tibial and sural nerve transection (TST) produce neuropathic pain behaviors, including spontaneous pain, tactile allodynia, and cold allodynia. Neuropathic pain behaviors in TST rats are not relieved by chemical sympathectomy using guanethidine, suggesting that this animal model may represent a SIP model.3 However, there are no reports on the effects of surgical lumbar sympathectomy on neuropathic pain behaviors of rats with TST. Therefore, we investigated the effects of surgical sympathectomy on pain behaviors in TST rats to determine whether TST model represents SIP-dominant neuropathic pain.
MATERIALS AND METHODS
Experiments were carried out on 12 male Sprague-Dawley rats, ranged from 180-220 g. Rats were housed in group cages on a 12-h night/day cycle with access to food and water at all times. Rooms were temperature- and humidity-controlled. The animal surgery and testing procedures were reviewed and approved by the Institutional Animal Care Committee at Yonsei University.
Rats were anesthetized using enflurane, after which a segment of the sciatic nerve was exposed between the mid-thigh level and the popliteal fossa. The three major divisions of the sciatic nerve (tibial, sural, and common peroneal nerves) were separated. The tibial and sural nerves were ligated using 4.0 black silk and transected, while the common peroneal nerve was left intact.3
Animals were divided into a sympathectomy group and a sham group. Seven days after the nerve transection, under anesthesia with a mixture of enflurane in O2, the lumbar sympathetic chain was located through a transperitoneal approach. In identifying levels of sympathetic ganglia, the most reliable landmark was found to be the left renal artery, which runs near the L2 ganglion. The L5 ganglion was usually located just rostral to the bifurcation of the descending aorta.4 Once each level was identified, the sympathetic chains along with ganglia on both sides were resected from L2 to L6 levels. For the sham group, the surgical procedures were the same except that the sympathetic chains and ganglia were not resected. Rats showing neurological motor deficits postoperatively were sacrificed immediately using thiopental sodium.
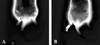
The success of the sympathectomy was confirmed by measuring skin temperature on the hind paw and by infra red thermography taken. The cutaneous temperature of the plantar hind paw surface was measured and infra red thermography was taken twice respectively: prior to surgical sympathectomy and 1 hour following the sympathectomy procedure. The temperature differences between the trunk and ipsilateral or contralateral hind paw were defined as ΔT1 or ΔT2, respectively.
Behavioral tests for mechanical and cold allodynia on the ipsilateral hindpaw were conducted on all rats 1 day prior to nerve injury, on days 2, 4 and 6 after nerve injury, and on days 1, 3, 7 and 14 after sympathectomy. Rats were placed in individual transparent acryl cages with wire mesh floors and allowed to adapt for 30 min.
For the tactile allodynia test, a mechanical threshold was measured by applying a von Frey hair (0.41-15.1 g) to the sensitive area (the medial aspect of the plantar surface) for 5 sec with pressure to slightly bend the hair. Brisk withdrawal or paw flinching was considered a positive response. In the absence of such a response, the filament of next greater force was applied. In the presence of a response, the filament of next lower force was applied. The cutoff value was defined as a stimulus intensity of 15 g. The 50% withdrawal threshold was determined using the up-down method.5
Cold allodynia was assessed using the acetone drop technique.6 A drop of acetone was applied to the sensitive area of the ipsilateral hindpaw, and each trial of acetone application was repeated five times at intervals of 3 min. Allodynia was assessed by measuring the frequency of foot withdrawal in response to acetone application. Rats with a mechanical threshold less than 3.0 g and a withdrawal in response to least 4 of the 5 acetone applications before sympathectomy (POD 6) were included in the study.
Data are presented as mean ± SEM. Statistical treatments were done using nonparametric statistical tests, including the Wilcoxon matched pairs and Friedman analysis of variance, followed by the Dunnett post hoc test. A p value of less than 0.05 was considered to be statistically significant.
RESULTS
The tactile threshold of all rats was about 15 g before nerve injury. This decreased dramatically by the 4th day after nerve injury (p < 0.05). There were no differences of threshold between the sympathectomy and the sham groups before sympathectomy. Surgical sympathectomy did not alter the tactile threshold in comparison with that measured one day before the sympathectomy, and there were no differences in tactile threshold between the two groups at any time point (Fig. 1A).
No rats responded to acetone application before nerve injury. The response frequency to acetone was significantly increased on the 2nd day after nerve injury (p < 0.05) with no differences of response frequency between two groups before the sympathectomy. Surgical sympathectomy did not significantly change the frequency of positive response to acetone compared with measurements taken one day prior to nerve injury. There were no differences in the response frequency between two groups at any time point (Fig. 1B).
Sympathectomy significantly increased skin temperature from 26.1 ± 0.1℃ to 26.7 ± 0.05℃ on the ipsilateral plantar surface and from 26.2 ± 0.05℃ to 26.65 ± 0.1℃ on the contralateral surface respectively (p < 0.05) (Fig. 2). Both ΔT1 (4.03 ± 0.51℃, 1.81 ± 0.21℃) and ΔT2 (4.15 ± 0.43℃, 2.66 ± 0.32℃) were significantly decreased after the sympathectomy compared with those before sympathectomy respectively (p < 0.05) (Fig. 3).
DISCUSSION
This experiment demonstrated that bilateral surgical sympathectomy did not improve existing tactile or cold allodynia after tibial and sural nerve transection.
Neuropathic pain resistant to conventional treatment has SMP and SIP portion.1 The most effective means of relieving SMP is to remove the sympathetic supply to the affected area. Sprouting of sympathetic postganglionic fibers that interact with the dorsal root ganglion and the proximal stump of the injured spinal nerve after peripheral nerve injury might be related to SMP.7 Sympathetic sprouting may vary with injury location or postoperative time course.8 The SNL model with the injury site closer to the DRG promotes a fast and pronounced sympathetic sprouting.9 Whether the sympathetic and afferent fibers interact in this model is uncertain. And if any interaction, the more distal nerve injury in TST model than in SNL might trigger a delayed or less sprouting of sympathetic fibers to the DRG. Therefore, early sympathectomy may have little effect on relieving neuropathic pain in this model, as the injury is more distal from the DRG than in any other neuropathic pain model.10,11
The SMP model has been studied extensively,2 but few models representing characteristic SIP have been introduced. The SNL model has been studied most extensively for features resembling SMP.12 Although the chronic constriction injury model and the partial sciatic nerve ligation model better represent SIP than SMP, some degree of sympathetic dependency has been shown in these two models.8 A Neuropathic pain model of sciatic cryoneurolysis was also introduced as a SIP-resembling model. However, this model shows incomplete neuropathic pain characteristics, with mechanical allodynia but no thermal hyperalgesia.13 In TST rats, spontaneous pain, mechanical and cold allodynia peak at 1-2 weeks and persist for 2-3 months. These are not improved by IP guanethidine,3,10 however, surgical sympathectomy has not been performed in the TST model. Although SMP is temporarily reversed by chemical sympathetic block, including phentolamine (blockade of α-adrenoceptors) or guanethidine (prevention of release of norepinephrine),2 there may be different results between the surgical sympathectomy and the chemical sympathetic block.14,15 The effectiveness of chemical sympathectomy may be dependent on the onset, duration, and the dosages of the administered drug.14,16 Hence, surgical sympathectomy may more reliable than a chemical sympathetic block. Bilateral sympathectomy was performed in our experiment because some of the lumbar sympathetic ganglia on both sides are often fused, making it difficult to perform a complete unilateral sympathectomy.4,12
The effectiveness of the sympathectomy may depend on the level of mechanical threshold and the time of sympathectomy. Sympathectomy is less effective in reducing mechanical hypersensitivity of the SNL model, especially when the mechanical threshold is high.12 In other words, surgical sympathectomy can reverse allodynia but not hyperalgesia.17 We could rule out the ineffectiveness of sympathectomy since the pre-sympathectomy mechanical threshold was low enough to be considered allodynia rather than hyperalgesia.
Lee et al. performed a chemical sympathectomy using IP guanethidine two weeks after TST.3 It may be difficult to know whether the changes in pain behaviors after the sympathectomy are a direct result of the sympathectomy or a simple reflection of behavioral signs of neuropathic pain, which may gradually decrease at 2 weeks. A Sympathectomy at a later stage would be of little value in determining whether TST would present as SIP. Therefore, our surgical sympathectomy was performed 1 week earlier than the chemical sympathectomy by Lee et al.3
The most common clinical response with a successful sympathectomy is an increase in perfusion to the distal part of the limb resulting in a significant increase in skin temperature.18 In our study, the increase of both ipsilateral and contralateral hindpaw skin temperature after bilateral sympathectomy suggests a lack of sympathetic innervation in the extremity after surgical resection of the paravertebral lumbar sympathetic chains. Evaluation of successful sympathectomy using the change of skin temperature might have some limitations. Whole body temperature might be reduced and changes in skin temperature may not be large enough to detect when internal organs are exposed to the ambient environment, and there may be a loss of thermoregulatory function during surgery. It will be more useful to compare the difference in temperature between the trunk and the extremities to assess increased peripheral blood flow after a sympathectomy. The infrared thermographic method provides an accurate and quantitative assessment of the effectiveness of sympathectomy.19
These data demonstrate that the majority of rats in the TST model exhibited characteristic neuropathic pain behaviors, including tactile and cold allodynia that peak approximately 1 week after nerve injury. TST-induced neuropathic pain behaviors are resistant to bilateral surgical sympathectomy, therefore the TST model can be a useful and easily reproducible model of sympathetically independent pain (SIP).


 XML Download
XML Download