

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Human telomerase is a ribonucleoprotein enzyme that is important for the maintenance of telomeric structures at the ends of chromosomes.1-3 Telomerase activity is closely associated with the expression of human telomerase reverse transcriptase (hTERT). The expression of hTERT RNA is detected at high levels in tumor tissues and tumor-derived cell lines but not in normal adjacent tissues or primary cells.4-6 A 5' flanking region of 341 bp in the hTERT gene was identified as the minimal promoter region, containing elements responsible for its activity.7,8 The differential activity of this region in normal cells and cancer cells opens the possibility of utilizing the hTERT promoter for the selective expression of apoptosis-inducing genes only in cancer cells.
TNF-related apoptosis inducing ligand (TRAIL) is a promising agent for development as a cancer therapeutic9 because it appears to specifically kill transformed and cancer cells, whereas most normal cells appear to be resistant to TRAIL.10,11 TRAIL induces apoptotic cell death upon binding to either of two pro-apoptotic TRAIL receptors, TRAIL R1 (DR4)12 or TRAIL R2 (KILLER/DR5).13 Although TRAIL has the potential to specifically kill cancer cells, many cancer cells are TRAIL-resistant.14,15
Therefore, we performed this study to examine whether the combination of TRAIL treatment and cancer cell-selective expression of TRAIL death receptor DR4 could induce cell death in TRAIL-resistant cancer cells. We generated an adenovirus expressing DR4 under the control of the hTERT promoter to express DR4 selectively in cancer cells. Here, we show that the hTERT promoter is active in a variety of human cancer cell lines but not in normal primary human keratinocytes, and further that the tumor-specific expression of DR4 induces apoptosis in TRAIL-resistant cancer cells when combined with TRAIL.
MATERIALS AND METHODS
Cell lines and culture
Normal human primary cervical epithelial cells that were kindly provided by Dr. Young-Tae Kim (Department of Obstetrics and Gynecology, Yonsei University College of Medicine) were grown in Keratinocyte-SFM medium (Gibco BRL, Rockville, MD, USA) containing bovine pituitary extract (40 µg/mL) and epidermal growth factor (1 ng/mL). Cancer cell lines were grown as previously described.14,16
Recombinant human TRAIL and caspase inhibitor treatments
Cells were treated with 100 ng/mL final concentration of TRAIL17 for 2 h. The irreversible pan caspase inhibitor z-VAD-fmk (R&D Systems, Minneapolis, MN, USA) was used at a final concentration of 20 µM and was added 2 h prior to the addition of TRAIL.
Cloning of the hTERT core promoter
The hTERT core promoter7 was amplified using genomic DNA as a template. For the amplification, the following primer pairs were used: Sense primer, 5'-GAAGATCT GGAGAGCTGCGCTGT-3' and anti-sense primer, 5'-GAAGATCTAAGCTT GCCAGGGCTT-3'. The amplified (301 bp) hTERT core promoter was cloned into the pGL3-basic (Promega, Madison, WI, USA) vector and was named pGL3-hTERT.
hTERT promoter activity assay
HCE4, CasKi, HCT116-neo, HCT116-E6, or normal human primary cervical epithelial cells were transiently transfected with pGL3-basic as a negative control, pGL3-control as a positive control, or pGL3-hTERT using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer's instructions. Briefly, 5 × 105 cells seeded in a 6-well plate were exposed to transfection mixtures containing 1.8 µg of the reporter plasmids and 0.2 µg of pCMV-β-galactosidase (BD Bioscience Clontech, Palo Alto, CA, USA) for 5 h at 37℃. Then, 3 mL of growth media were added to the cells, followed incubation for an additional 16 h. The cells were harvested 48 h after transfection. A luciferase ssay was performed according to the manufacturer's protocol (Promega). A β-galactosidase assay was performed to standardize transfection efficiency. In cells infected with adenovirus, infections were carried out 24 h after transfection.
Generation of adenoviruses
To make an adenovirus expressing full length DR4 under the control of the hTERT core promoter, the pShuttle-CMV (Stratagene, La Jolla, CA, USA) vector was used. To obtain DR4 expression driven by the hTERT core promoter, the CMV promoter in pShuttle-CMV was replaced by a PCR amplified hTERT core promoter (pShuttle-hTERT) and then full length DR4 cDNA was inserted into the pShuttle-hTERT (pShuttle-hTERT-DR4) vector. Recombinant plasmid was generated in BJ5183 E. coli by co-transforming PmeI-digested pShuttle-hTERT-DR4 and pAdEASY.18 The recombinant plasmid was digested with PacI and was used for transfection into 293 cells using Lipofectamine 2000 (Invitrogen). Approximately 7 days after transfection, the cells were harvested, and the viruses were amplified and purified as previously described.19 A TRAIL-expressing adenovirus was made using the extracellular portion of TRAIL (Aa 95 to 281), which can be secreted and self-oligomerized. A DNA fragment that encodes the secretion signal from the V-J2-C region of the mouse Ig κ chain signal peptide, isoleucine zipper (ILZ) fragment that was made by annealing two synthetic oligonucleotides (5'-GGGGTACCATGAAGCAGATCGAGGACAAAATT GAGGAAATCCTGTCCAAGATTTACCACATCGAGAACGAGATCGCCCGGATTAAGAAACTCATTGGCGAGAAGCTTGGG-3' and 5'-CCCAAGCTTCTCGCCAATGAGTTTCTTAATCCGGGCGATCTCGTTCTCGATGTGGTAAATCTTGGACAGGATTTCCTCAATTTTGTCCTCGATCTGCTTCATGGTACCCC-3') for self-oligomerization20 and was sequentially inserted into the pAdTRACK/CMV vector. An adenovirus expressing the E6 ORF of human papillomavirus (HPV) type 16 was generated (Ad-E6). An adenovirus expressing green fluorescence protein (GFP) was also generated and used as a control virus (Ad-GFP). The infectivity of viruses was checked by the observation of green fluorescence within infected cells using fluorescence microscopy. To confirm the oligomerization of TRAIL, ethylene glycol bis-(succinic acid) (EGS, final 1 mM; Sigma, St. Louis, MO, U.S.A.) was used as a cross-linker to maintain oligomeric structure during denaturing SDS-PAGE analysis.
Western blot immunostaining
Western blot analysis was carried out as previously described.14 Blotted membranes were immunostained with anti-DR4 (1:500, Pharmingen, San Diego, CA, U.S.A.), anti-caspase-3 (E-8, 1:200; Santa Cruz, Santa Cruz, CA, U.S.A.), anti-PARP (1: 1,000; Cell Signaling Technology, Inc., Beverly, MA, U.S.A.), anti-TRAIL (1:500; PeproTech Inc., Rocky Hill, NJ, U.S.A.), or anti-α-tubulin (1:500; Oncogene Science, Cambridge, MA, U.S.A.).
Flow cytometric cell death analysis
Cells were harvested after the indicated treatments and time points, stained with propidium iodide, and analyzed by flow cytometry for sub-G1 content as previously described.21 Cell sorting was performed on a BD FACSCalibur (Becton Dickinson, Boston, MA, U.S.A.).
RESULTS
hTERT core promoter activity in normal and cancer cell lines
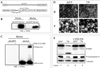
To examine the promoter activity of the hTERT gene in normal human cervical keratinocytes or in a TRAIL-resistant human esophageal cancer cell line, HCE4, a luciferase activity assay was performed. Transfection of pGL3-hTERT into HCE4 cells yielded a significant increase in promoter activity compared to the transfection of the pGL3 basic control vector (Fig. 1B). Transfection of the same construct into normal keratinocytes resulted only in background levels of luciferase activity (Fig. 1A). The E6 protein from HPV type 16 transcriptionally transactivates the hTERT promoter;1 expectedly, hTERT promoter activity was markedly increased in Ad-E6 infected cells (Fig. 1). These results suggest that the hTERT promoter is active only in cancer cells and that the presence f E6 protein could augment hTERT promoter activity.
Expression and secretion of TRAIL by TRAIL-expressing adenovirus
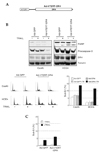
We generated an adenovirus expressing the isoleucine zipper (ILZ)-fused extracellular portion of TRAIL (Ad-ILZ-TRAIL, Fig. 2A). To confirm the expression, secretion, and oligomerization of TRAIL, TRAIL-resistant HCE4 cells were infected with Ad-ILZ-TRAIL. Two days after infection, Western blot analysis was performed using cell extracts and the media. In the cell extracts, both un-processed and processed forms of TRAIL were present, but in the media only the processed form of the expressed TRAIL was present (Fig. 2B). The oligomerization of the secreted TRAIL was confirmed by cross-linking followed by Western blot analysis (Fig. 2C). The biological activity of TRAIL was examined using TRAIL-sensitive SW480 colon cancer cells. The cells infected with Ad-ILZ-TRAIL were morphologically apoptotic (Fig. 2D, TR). Procaspase-3 was activated and PARP was cleaved in the cells infected with Ad- ILZ-TRAIL but not in the presence of z-VAD-FMK (Fig. 2E. lane 2 vs. 4). Together, these data suggest that TRAIL expressed from the adenovirus is secreted, oligomerized, and biologically active.
Cancer cell death induced by the combination of TRAIL treatment and Ad-hTERT-DR4 infection
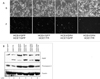
An adenovirus that expresses DR4 under the control of the hTERT core promoter was generated (Fig. 3A). When infected into HCE4 or CasKi cells, the expression level of DR4 was slightly increased and the cells underwent apoptosis upon TRAIL treatment (Fig. 3B). Infection of normal primary human cervical epithelial cells with either Ad-GFP or Ad-hTERT-DR4 did not result in cell death. The addition of TRAIL to infected cells slightly increased cell death, but less than 10% of the cells underwent apoptosis (Fig. 3C). Taken together, these results suggest that the combination of TRAIL treatment and the infection of an adenovirus expressing TRAIL-death receptor DR4 under the control of the hTERT core promoter has the potential to induce cell death in TRAIL-resistant cancer cells but not in normal cells.
The bystander effect of TRAIL from Ad-ILZ-TR
To check the bystander effect of TRAIL produced from the cells infected with Ad-ILZ-TRAIL, TRAIL-resistant HCE4 cells were infected with either Ad-GFP or Ad-hTERT DR4, and TRAIL-resistant HCE7 cells16 were infected with either Ad-GFP or Ad-ILZ-TRAIL. One day after the infection, the cells were harvested and co-cultured for 24 h. When Ad-hTERT DR4 infected HCE4 cells and Ad-ILZ-TRAIL infected HCE7 cells were co-cultured, the induction of apoptosis was evident both morphologically (Fig. 4A) and biochemically (Fig. 4B). These results suggest that DR4-expressing cancer cells might be killed by the bystander effect of TRAIL encoded by the Ad-ILZ-TRAIL vector.
DISCUSSION
Although TRAIL is a potent inducer of apoptosis in cancer cells, there are still several TRAIL-resistant cancer cells. Many successful approaches have been devised to overcome this resistance, such as the combination of TRAIL with other reagents like DNA damaging agents,14,22 ionizing radiation,23 or virus expressing wild type p53.24 In this study, we tested the effect of combining TRAIL and hTERT core promoter-driven expression of DR4 on the induction of apoptosis in TRAIL-resistant cancer cell lines and normal cells. We found that the hTERT core promoter was able to drive DR4 expression in cancer cells. The combination of infection with a DR4-expressing adenovirus and TRAIL treatment was able induce apoptosis in TRAIL-resistant cancer cells without inducing significant cell death in primary normal cells. In addition, TRAIL expressed from an adenovirus could induce cell death in Ad-hTERT-DR4 infected TRAIL-resistant cancer cells by the bystander effect.
The hTERT promoter was widely used for inducing apoptosis selectively in cancer cells by driving expression of apoptosis-inducing gene products such as bax or TRAIL.25-30 However, the expression of TRAIL alone could only induce apoptosis in TRAIL-sensitive cancer cells and the expression of bax alone could induce apoptosis only in cancer cells that were infected with the adenoviruses. Accordingly, the combined use of the adenovirus that could express TRAIL-death receptor selectively in cancer cells and the adenovirus that could express TRAIL which is then secreted into adjacent cancer cells, thereby exerting the bystander effect, may induce apoptosis in clumps of cancer cells that are resistant to TRAIL. Thus, the generation of an adenovirus that could bicistronically express the death receptor and TRAIL under the control of the hTERT promoter could reduce the number of viral particles used in the experiment, thereby alleviating adenoviral toxicity toward normal cells.
The strategy introduced in this experiment for inducing cancer cell death could be applied to the treatment of cervical cancer because many cervical cancers are associated with the infection of oncogenic HPV, such as types 16 and 18, which can transform normal cells by expressing E6 and E7 oncoproteins.

 XML Download
XML Download