

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
In this study brain astrocytes were examined from the standpoint of their cytotoxic effects. During inflammation, significant astrocyte death occurs after a period of reactive gliosis. Such gliosis has been reported in Alzheimer's disease,1 multiple sclerosis2,3 and HIV infections.4 TNF-related apoptosis inducing ligand (TRAIL) is a member of the death ligand/receptor pairs, and acts independently of Fas mediated apoptosis.5 TRAIL is consistently expressed in low-grade astrocytomas, anaplastic astrocytomas, and glioblastomas.6-8 However, little published information is available about the relationship between primary astrocytes and TRAIL, though we previously issued a report which demonstrated that the mRNA of TRAIL is detectable in primary astrocytes after stimulation with pro-inflammatory cytokines.9 When inflammation, tumor progression, or a degenerative disease occurs in the CNS, lymphocytes infiltrate and secrete IFN-γ, priming the immune reactions of astrocytes. We previously reported that the full transcriptional activity of STAT1 is manifested when the Ser727 is phosphorylated, and that this is dependent on p38 MAP kinase in astrocytes.10 Here, the expression of TRAIL in primary astrocytes induced by IFN-γ, and TRAIL-mediated apoptosis were addressed in an attempt to identify the functional role of astrocytes in the apoptosis of brain cells.
MATERIALS AND METHODS
Preparation of human brain astrocytes
Fetal astrocytes were isolated from a 25-week-old human fetus. The brain samples were obtained by therapeutic abortion. The use of tissue samples was approved by the Institutional Review Board. Astrocytes were cultured in 10% FCS (Gibco BRL, Grand Island, NY)-DMEM (Gibco BRL) containing 1% nonessential amino acids (Sigma, St. Louis, MO) at 37℃ in 5% CO2. Primary cultures were maintained for up to ten weeks. Staining for glial fibrillary acidic protein (GFAP) was done using anti-GFAP antibody (Pharmingen, San Diego, CA) to estimate the pure astrocyte population, which revealed approximately 99% of the cells were astrocytes. GFAP is the major component of the intermediate filaments of astrocytes and is commonly used as an astrocyte marker. Staining with anti-HLA-DR antibody (Becton Dickson, Franklin Lakes, NJ) was done to detect microglias and activated astrocytes. Anti-GFAP or anti-HLA-DR antibodies were conjugated with FITC and used as recommended by the manufacture.
RNase protection assay (RPA), RT-PCR and southern blotting
For the induction of TRAIL, astrocytes were cultured with IFN-℃ (Genzyme, Cambridge, MA) at a concentration of 250 U/mL or 500 U/mL for 8 or 24 hr. IL-1β (Genzyme) was treated at 100 U/mL, and TNF-α (Genzyme) was treated at 750 U/mL for 8 hr for RNA preparation.
Total RNA was isolated using an RNeasy kit (Qiagen, Santa Claris, CA). The level of TRAIL mRNA was measured by RPA or RT-PCR. For RPA, a RiboQuant multiprobe RNase protection assay kit (Pharmingen) was used with 10 µg of total RNA. For RT-PCR, total RNA (1 µg) was used to synthesize cDNA using 0.1 O.D. of random hexamer (Pharmacia, Uppsala, Sweden) and 200 U of M-MLV reverse transcriptase (Gibco BRL). The TRAIL primers used were; forward: 5'-CCCAATGACGAAGAGAGTATGA-3'; reverse: 5'-GGAATAGATGTAGTAAAACCCT-3', and the PCR conditions were as follows: denaturation at 94℃ for 30 sec, annealing at 59℃ for 30 sec and extension at 72℃ for 30 sec. The PCR buffer contained 10 mM Tris-HCl (pH 10), 2.0 mM MgCl2, 50 mM KCl with 1.25 U of Taq polymerase (Takara, Tokyo, Japan). After 30 PCR cycles, an additional extension at 72℃ for 10 min was performed for all primers. β-actin was used as the internal control. Southern blotting was performed using the TRAIL PCR product (100 ng) labeled using an ECL kit (Amersham life Science, Piscataway, NJ) and hybridized at 42℃ overnight.
Apoptosis assay
To evaluate the function of the induced TRAIL, astrocytes pre-treated with IFN-γ for 24 hr were reacted with Peer cells, which are sensitive to TRAIL-mediated apoptosis.11 Peer cells are of lymphoid origin, and were maintained in suspension culture at 10% FCS-RPMI. Adherent astrocytes (effector cells) and floating Peer cells (target cells) were reacted at E:T ratios of 1:1, 2:1, or 4:1. Reactions were performed in 10% RPMI 1640 for 24 hr. Apoptotic cells were stained using Annexin V-FITC (Trevigen, Gaithersburg, MD) and dead cells using propidium iodide (Sigma, St. Louis, MO). 1 × 105 cells were analyzed using a FACstar (Becton Dickinson, San Jose, CA).
Western blotting
For western blotting, each protein (40 µg) of each whole cell homogenate was loaded on 12% PAGE. Proteins were blotted onto a nitrocellulose membrane (Amersham Pharmacia Biotech., Piscataway, NJ) and incubated with 5 µg/ml of polyclonal mouse anti-human TRAIL antibody (Pharmingen). After three washes in PBS containing 0.1% Tween 20, the membranes were incubated with secondary peroxidase-conjugated goat anti-mouse IgG (Jackson Immuno Research, Baltimore, MD) for 1 hr, washed three times in PBS containing 0.1% Tween 20, and developed using an ECL system (Amersham Pharmacia Biotech).
RESULTS
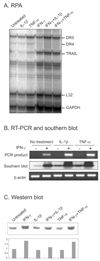
To evaluate TRAIL expression RPA was done after treating brain astrocytes with IFN-γ, IL-1, or TNF-α. All of these cytokines induced TRAIL expression, but the effect of IFN-γ was prominent (Fig. 1A). RT-PCR and the subsequent southern blot analysis of the RT-PCR product revealed that TRAIL mRNA was present at a very low level (Fig. 1B), and that IFN-γ increased the level of TRAIL RNA. Western blotting showed TRAIL protein was also increased by IFN-γ (Fig. 1C).
To determine the biological function of the expressed TRAIL, primary astrocytes treated with IFN-γ for 24 hr were reacted with Peer cells, which have been reported to be TRAIL-sensitive target cells. After reaction with astrocytes (effector cells), floating Peer cells (target cells) were collected carefully; the astrocytes remained adherent. The Peer cells were then stained with annexin V-FITC and propidium iodide. The percentages of annexin V positive cells in the target Peer cell population were 26.0% (E:T = 1:1), 41.4% (E:T = 2:1) or 43.9% (E:T = 4:1) after reaction with IFN-γ treated astrocytes as compared with 3.9% (E:T = 1:1), 5.3% (E:T = 2:1), or 6.2% (E:T = 4:1) for IFN-γ untreated astrocytes (Fig. 2). These results suggest that induced TRAIL molecules on astrocytes are functionally active and trigger apoptosis in proper target cells. Treatment with IFN-γ for 24 hr did not induce any apoptotic changes in primary astrocytes themselves (data not shown).
DISCUSSION
Astrocytes have been reported to play a significant role in the immune responses of the CNS.12 They produce various cytokines 13 and express the cell death mediated molecules Fas and FasL.3,9 Here, we report that the expression of TRAIL can be induced in primary astrocytes by IFN-γ treatment. In this study, human astrocytes expressed TRAIL at very low levels during primary culture, however, IL-1β, TNF-α, and more significantly IFN-γ up-regulated TRAIL expression. However, it has been suggested that the human brain lacks TRAIL expression,14 this difference may reflect the in vivo status of TRAIL expression. The low level of TRAIL expression detected in control astrocytes in our study may have been due to the slight activation of astrocytes during culture. However, we observed TRAIL expression after IFN-γ treatment by western blotting. In line with the results of the present study, a recent report showed that TNF and TRAIL do not induce apoptosis in reactive astrocytes.15 IFN-γ has been shown to increase the expression of the TRAIL in human colon cancer cell lines16 and in human monocytes.17 Moreover, the up-regulation of TRAIL expression has been reported in NK cells of the liver, spleen, and lungs, by the administration of IL-12, a powerful inducer of IFN-γ production in NK cells and NKT cells.18 Moreover, TRAIL expression was not detected in the NK cells of IFN-γ knock out mice.18
In conclusion, the following were found in human brain astrocytes; 1) TRAIL was expressed at a very low level and this expression was increased by IFN-γ stimulation, 2) astrocytes can induce TRAIL-mediated apoptosis when they were reacted with appropriate target cells. Moreover, the up-regulation of TRAIL may contribute to immune privilege during fetal brain development and to the apoptosis of neighboring cells during pathological conditions.

 XML Download
XML Download