

 PDF
PDF ePub
ePub Citation
Citation Print
Print


INTRODUCTION
Osteoporosis and other diseases involving bone loss are a major public health problem. Despite the recent successes with drugs that inhibit bone resorption, notably bisphosphonates, there is a clear therapeutic need for bone anabolic molecules, particularly in patients who have already suffered substantial bone loss.
Parathyroid hormone (PTH) and prostaglandin E2 (PGE2) stimulate bone formation in experimental animals and humans.1-3 Several studies suggest that cyclic AMP (cAMP), which initiates protein kinase A (PKA) signaling, mediates the anabolic effects of these two molecules.3,4 Many cAMP-responsive genes have been identified in PTH-treated osteoblasts, including collagenase,5 c-fos,6 type I collagen,7 interleukin-6,8 cycloxygenase-2 (cox-2),9 TNF-related activation-induced cytokine (TRANCE, also known as RANKL, ODF, or OPGL),10 and inducible cAMP early repressor (ICER).11
ICER is a member of the cAMP response element binding protein (CREB) and CRE modulator (CREM) family of transcription factors, which bind to CREs.12 The ICER is generated in an inducible manner when an internal promoter of the CREM gene, containing CRE sites, is stimulated by increased cAMP levels.12 Because the ICER consists of only a DNA-binding domain identical to the one in the CREM and lacks the transactivation domain, the ICER serves as a dominant-negative of CREM/CREB-mediated transcription.12
Intracellular cAMP is generated by adenylate cyclase from adenosine triphosphate (ATP) as a substrate, whereas cAMP-specific phosphodiesterases (PDEs) catalyze the hydrolysis of cAMP to 5'-AMP.13,14 Therefore, the intracellular cAMP gradients are governed by a balance between its generation by adenylate cyclase and degradation by the PDEs.
The PDE family consists of 11 isozymes ranging from PDE1 to 11. Those isozymes involved in the degradation of cAMP are PDE1, 2, 3, 4, 7, 8, 10, and 11, with some of these PDE isozymes being further classified into subtypes.14 Rolipram, a PDE4 specific inhibitor, has recently been demonstrated to increase the bone mass mainly by promoting bone formation in normal mice.15 Furthermore, PDE4 inhibitors have been shown to have therapeutic effects in different experimental osteopenia models.16,17 Although it has been hypothesized that PDE4 inhibitors can mimic the anabolic effects of PTH and PGE2 on the bone, little is known about the precise mechanism by which the PDE4 inhibitors regulate the expression of the osteoblastic genes.
In this study, rolipram was shown to induce ICER mRNA expression in mouse osteoblastic cells. It was found that rolipram-dependent ICER mRNA expression was mediated possibly by the PKA and p38 mitogen-activated protein kinase (MAPK) pathway, with little contribution from the extracellular signal-regulated kinase (ERK) MAPK pathway. It was also suggested that ICER might play an important modulatory role in the rolipram-mediated regulation of TRANCE, which is an essential molecule for osteoclastogenesis,18-20 in osteoblasts.
MATERIALS AND METHODS
Reagents
H89, PD98059 and SB203580 were obtained from Calbiochem (San Diego, CA). All other reagents were purchased from Sigma-Aldrich (St. Louis, MO).
Cells
Primary calvarial osteoblasts were isolated from the calvariae of neonatal ddY mice (Japan SLC Inc., Shizuoka, Japan) by a conventional method using 0.1% collagenase and 0.2% dispase. UAMS-32, which is an osteoblastic/stromal cell line, was a kind gift from Prof. Masamichi Takami (Showa University, Tokyo, Japan). All the cells were cultured in α-MEM/10% FBS at 37℃ and 5% CO2.
RT-PCR analysis
Total RNA (1 µg) was reverse-transcribed using Superscript II (Invitrogen, CA, USA) according to the manufacturer's protocols. Aliquots of the obtained cDNA pool were subjected to PCR amplification with Go Taq DNA polymerase (Promega Co., WI, USA). The primers for ICER and glyceraldehydes-3-phosphate dehydrogenase (GAPDH) used in this study are as follows: ICER, 5'-gatactggagatgaaactga-3' (forward), 5'-ctttctcatacagttcacag-3' (reverse); and GAPDH, 5'-gaaggtcggtgtgaacggatttggc-3' (forward), 5'-catgtaggccatgaggtccaccac-3' (reverse). The PCR program is as follows: 40 (ICER) or 28 (GAPDH) cycles, after an initial denaturation step at 94℃ for 3 minutes, then denaturation at 94℃ for 30 seconds, annealing at 48℃ (ICER) or 52℃ (GAPDH) for 45 seconds, and extension at 72℃ for 60 seconds, with a final extension at 72℃ for 10 minutes.
Immunoblot analysis
Total protein extracts were isolated from the rolipram-treated UAMS-32 cells. After separation in SDS-PAGE, the proteins were transferred onto Immobilon-P membranes (Millipore, Bedford, MA). The membranes were blocked with 5% nonfat-milk in TBS-T (150 mM NaCl, 20 mM Tris, pH 7.4, 0.1% Tween 20), and then immunostained with either the anti-phospho p38 antibody (1:1000) or the anti-p38 antibody (1:1000) followed by secondary horseradish peroxidase-conjugated antibody (1:5000, Cell Signaling Technology). The membranes were developed using an enhanced chemiluminescence detection kit (Amersham Biosciences, Bukinghamshire, UK).
Northern blot analysis
Cells in 60-mm diameter dishes were cultured with the agents for the indicated periods, and subjected to total RNA isolation using Trizol reagent (Invitrogen, CA, USA) according to the manufacturer's protocol. The total RNA (20 µg) was electrophoresed in 1.2% agarose-formaldehyde gels, transferred to nylon membrane filters (Hybond N+, Amersham Biosciences, Bukinghamshire, UK), and hybridized with the 32P-labeled cDNA probes. The cDNAs encoding TRANCE and GAPDH cloned by polymerase chain reaction (PCR) were used as the probes. After the final wash, the membrane was exposed to an X-ray film (BioMax, Kodak, Rochester, NY) at -70℃.
RESULTS
Rolipram induces ICER mRNA expression in osteoblastic cells
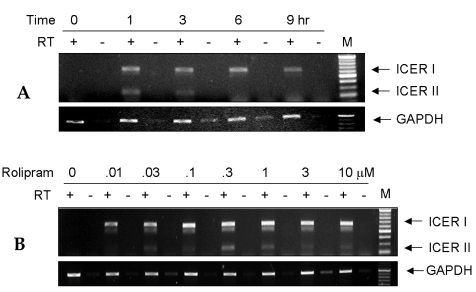
The effect of rolipram, a selective PDE4 inhibitor, on ICER mRNA expression was examined using RT-PCR in osteoblastic cells. Although there are four isozymes in the ICER family (ICER-I, ICER-Iγ, ICER-II, and ICER IIγ), the function of each isoform identified thus far is indistinguishable.12 Rolipram induced mRNA expression of ICER-I (700 bp) and ICER-II (262 bp) in UAMS-32 cells as early as 1 hr, which persisted for up to 9 hrs (Fig. 1A). In order to determine the dose-dependence of rolipram-induced ICER mRNA expression, the UAMS-32 cells were treated with various concentrations of rolipram for 3 hrs. Fig. 1B shows that 0.01-10 µM rolipram stimulated the ICER in a dose-dependent manner. These results demonstrate that rolipram strongly induces ICER mRNA expression in osteoblastic cells.
Inhibitors of PKA and p38 MAPK, but not of ERK, suppress ICER mRNA expression induced by rolipram
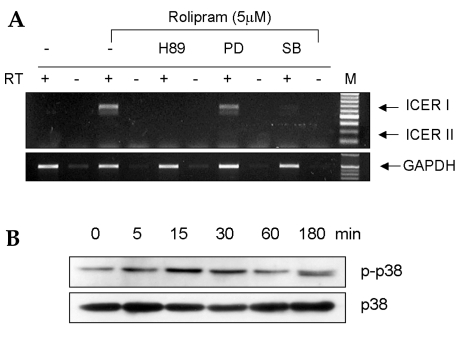
In order to identify the signaling pathways used by rolipram to induce ICER expression, the roles of the PKA-, ERK-, and p38 MAPK signaling cascades in ICER expression were examined. The cAMP-PKA pathway is essential for PTH-dependent ICER expression in osteoblasts.21 Consistent with this, pretreatment of UAMS-32 cells with H89, a PKA inhibitor, decreased the ICER mRNA expression levels induced by rolipram (Fig. 2A). Furthermore, SB203580, a p38 MAPK inhibitor, also decreased the ICER mRNA expression levels induced by rolipram. However, a MAPK/ERK kinase (MEK) inhibitor, PD98059, did not affect the ICER mRNA levels induced by rolipram (Fig. 2A). Immunoblot analysis with a phospho-p38-specific antibody confirmed that rolipram stimulates the rapid phosphorylation of p38 with its maximum effect at 15 min (Fig. 2B). This suggests that rolipram regulates ICER mRNA expression in osteoblastic cells possibly via the activation of the PKA and p38 MAPK signaling pathways, but not via the activation of the ERK MAPK pathway.
ICER may play an important modulatory role in attenuating TRANCE mRNA expression in osteoblasts
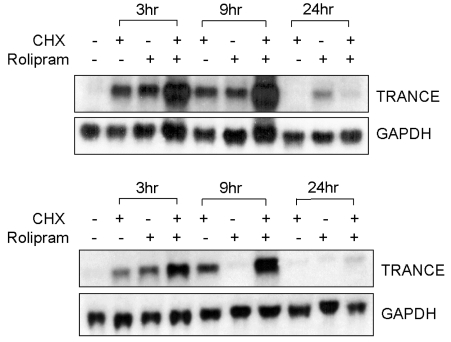
It was previously reported that rolipram stimulates TRANCE mRNA expression in osteoblastic cells.22 In order to determine if intermediate gene expression is essential for the effects of rolipram on TRANCE expression, the UAMS-32 cells were treated with rolipram for 3, 9, 24 hrs in the presence or absence of the protein synthesis inhibitor, cycloheximide. Cycloheximide alone stimulated the TRANCE mRNA levels at 3 and 9 hrs but did not block the additional stimulatory action of rolipram in the UAMS-32 cells (Fig. 3A). This indicates that rolipram stimulates TRANCE mRNA expression in osteoblastic cells directly. The expression level of TRANCE mRNA by rolipram decreased at 9 hrs and returned almost to the baseline by 24 hrs (Fig. 3A). However, pretreatment of cycloheximide caused a continuing increase in the TRANCE mRNA expression level at 9 hrs, suggesting the possible involvement of a transcriptional repressor such as ICER on TRANCE expression in osteoblastic cells. Primary mouse calvarial cells were used in order to exclude the possibility that the effect of cycloheximide on rolipram-induced TRANCE expression is unique in UAMS-32 cells. As shown in Fig. 3B, similar results were also obtained in primary mouse calvarial cells.
DISCUSSION
This study showd that rolipram, a PDE4 inhibitor, rapidly induces ICER mRNA in UAMS-32 osteoblastic cells (Fig. 1). A similar induction of ICER mRNA by cAMP was reported in other cells and tissues including pituitary corticotrophs,23 PC12 cells,12 mouse calvaria11 and rat thyroid gland.24 The stimulation of the PKA pathway has been suggested to be a mediator of cAMP-induced ICER mRNA expression in many experimental models.12,24,25 In agreement with this, rolipram was found to induce ICER mRNA expression in osteoblastic cells via the PKA signaling pathway (Fig. 2A).
Interestingly, nerve growth factor (NGF) was demonstrated to induce ICER in adrenal PC12 cells via the ras-MAP kinase pathway.26 It was shown that the activation of p38 MAPK, as well as PKA, possibly mediates ICER mRNA induction by rolipram in osteoblastic cells (Fig. 2). These results suggest that ICER expression may not be exclusively coupled to the cAMP-PKA pathway.
ICER is believed to be a repressor of cAMPinduced transcription. For example, in the pineal gland ICER has been reported to be involved in the repression of melatonin synthesis during the course of the normal circadian rhythm.25 ICER inhibits the PKA-stimulated transcriptional activity of tyrosine hydroxylase in PC12 cells.27 ICER has also been shown to play a regulatory role in PTH-stimulated COX-2 transcription in osteoblastic cells.9 The mechanism by which ICER inhibits the cAMP-inducible genes typically includes binding to a variety of cAMP response elements (CREs) or CRE-like sequences in the promoters of target genes that are binding sites for the activator forms of the CRE-binding protein (CREB) and/or the related factors of the CREM.12 Because the ICER lacks the CREM transactivation domain,12 which is similar in sequence to that of CREB, the binding of ICER to a CRE does not allow the recruitment of either CREB or the CREB-binding protein (CBP/p300) to activate transcription.
The activation of the cAMP-PKA pathway induces the expression of TRANCE in osteoblasts, which triggers osteoclast differentiation.28,29 Recently, cAMP-stimulated TRANCE expression was demonstrated to involve CREB-mediated transcription.30 The attenuation of rolipram-induced TRANCE mRNA expression was shown to require the de novo protein synthesis in osteoblasts (Fig. 3), suggesting the possible role of a repressor protein such as ICER. It has been suggested that the rapid activation of ICER could serve as a protective mechanism by which the cell limits mRNA accumulation of the CRE-containing genes, and that this regulation subverts the pathological consequences of the chronic stimulation of activation of PKA.23 Therefore, the enhanced expression of TRANCE by the activation of PKA would be determined by a complex balance of activation by CREB and repression by ICER, providing a system for the fine modulation of gene expression. Although no direct evidence on the functional ability of ICER to negatively regulate TRANCE mRNA expression is currently available, it is likely that future studies will reveal the additional roles of the ICER in the down-regulation of TRANCE mRNA expression.
 XML Download
XML Download